Восстановление окружения молекулы белка в кристалле
ДНК-белковый комплекс 3HDD
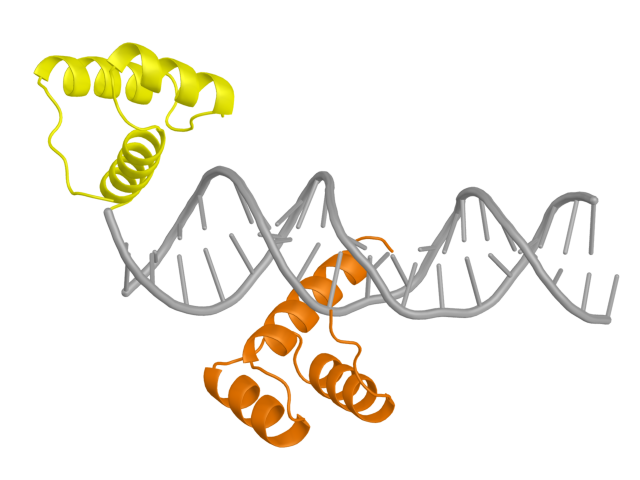
Из банка PDB была скачана структура ДНК-белкового комплекса 3HDD (рис. 1). Две цепи белка были покрашены а разные цвета (желтый и оранжевый на картинке ниже), ДНК обозначена серым цветом. Видно, что цепь B находится на самом конце ДНК, и пока что не очень понятно, чем это обусловлено.Рис.1: Структура 3HDD
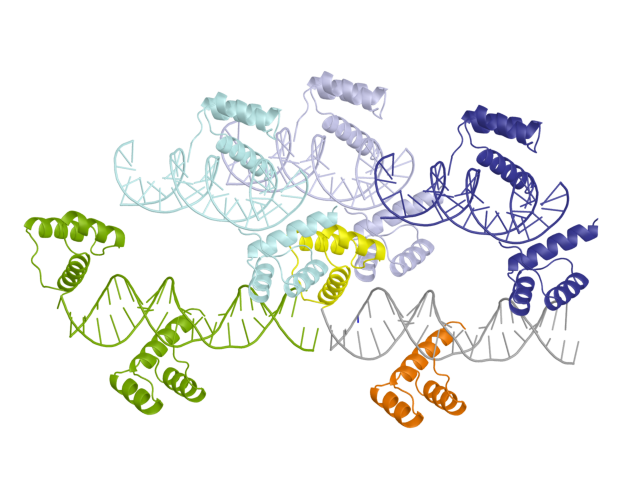
Командой "symexp near, 3HDD, chain B, 4" были восстановлены соседние с этой цепью кристаллографические ячейки, расположенные на расстоянии не более 4 А от нее (так мы точно увидим все водородные связи этой цепи). Разными цветами (зеленый, синий, бледно-голубой и бледно-фиолетовый).
Рис.2: Структура 3HDD с восстановленными соседними кристаллографическими ячейками
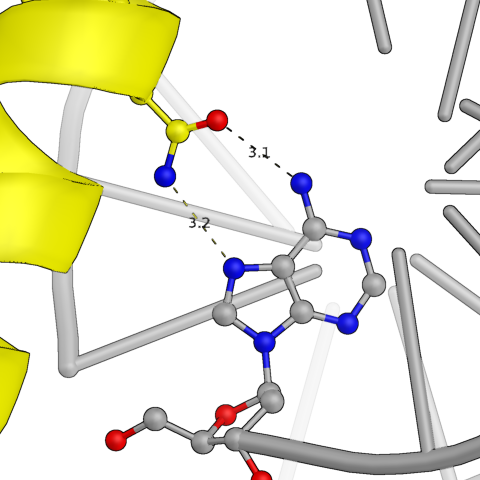
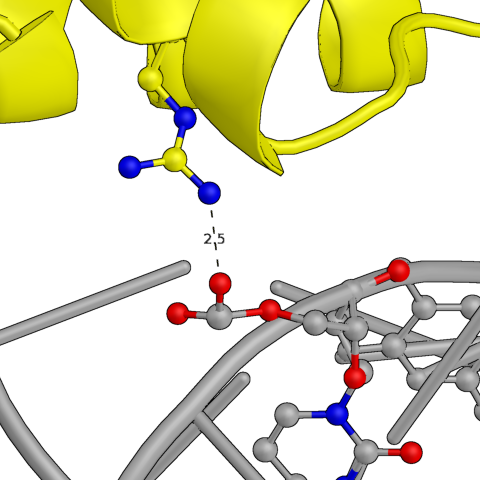
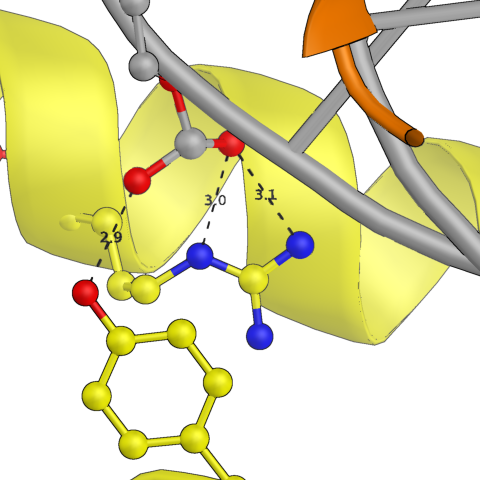
Сначала были проанализированы водородные контакты между белковой цепью и той ДНК, которая была в исходной ячейке. На картинках ниже представлены некоторые из них. По большей части боковые радикалы белка взаимодействуют с фосфатами ДНК, но есть и связи непосредственно с азотистым основанием. Участвуют в связи как аминокислотные остатки альфа-спиралей, так и петель.
Рис.3: Примеры водородных связей между белком и цепью ДНК из исходной ячейки
| Asn — Adenine (base) | Arg — Cytosine (phosphate) | Arg, Tyr — Thymine (phosphate) |
 |  |  |
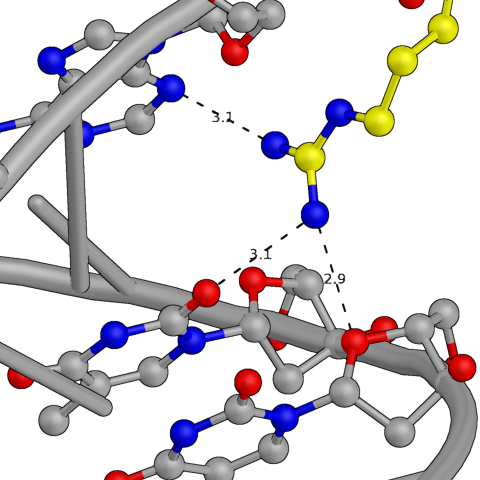
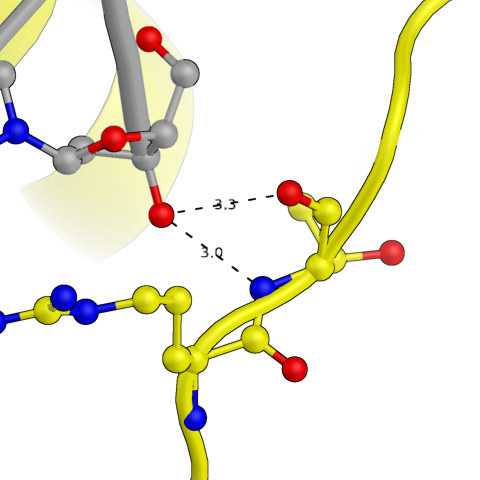
Затем были изучены водородные связи между той же белковой цепью и одной из ДНК из соседней восстановленной ячейки (покрашена в зеленый на общей картинке). Эта ДНК расположена непосредственно как продолжение исходной. Вероятно, это обусловлено стекинг-взаимодействиями между молекулами ДНК, а так же связями с этой самой белковой цепью.
Рис.4: Примеры водородных связей между белком и цепью ДНК из соседней ячейки
| Arg — Adenine (base) + Thymines (base, sugar) | Thr — Adenine (phosphate) | Lys — Adenine (phosphate) |
 |  | 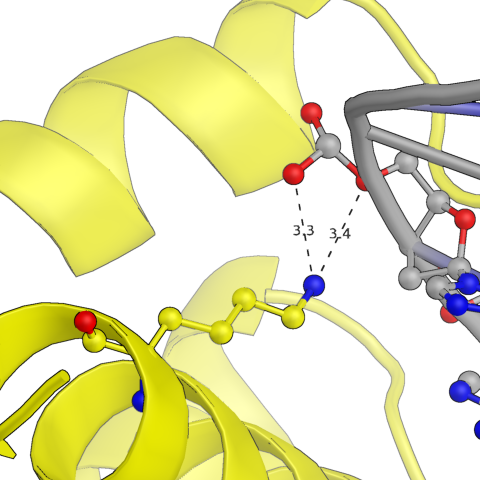 |
С другой стороны белковой цепи она взаимодействует с ДНК из другой кристоллаграфической ячейки (покрашена темно-синим), причем возможные водородные связи образуются глутамином и аргинином, сориентированными в одну сторону относительно альфа-спирали. В целом взаимодействий здесь намного меньше, чем с первыми двумя нуклеиновыми кислотами.
Рис.5: Примеры водородных связей между белком и цепью ДНК из соседней ячейки
Gln, Arg — Guanine, Cytosine (phosphates)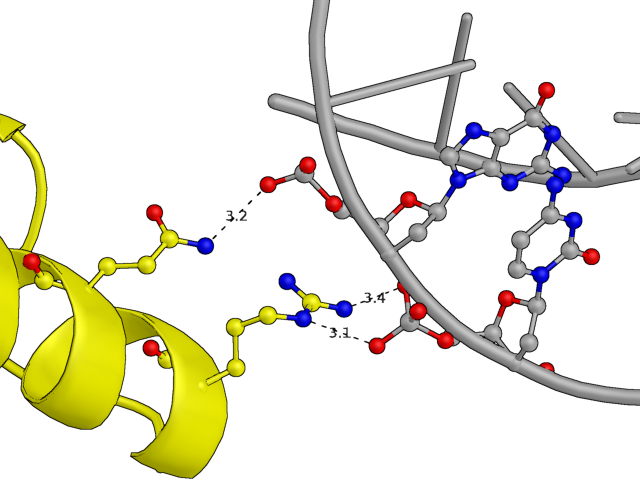
У ДНК из третьей и четвертой кристаллографических ячеек (покрашены в бледные оттенки) взаимодействий с белком найдено не было. И правда, там скорее взаимодействуют между собой белковые цепи, а ДНК расположена далеко от исходной ячейки.
Вывод:
Странное на первый взгляд расположение одной из цепей белка в структуре объясняется тем, что на самом деле в кристалле присутствуют и другие молекулы ДНК и белков. Расположенные в соседних ячейках, они не видны в исходной структуре, но взаимодействуют с ней. Конкретно для этой структуре были найдены еще две ячейки, взаимодействие с ДНК которых стабилизирует белок именно в этом положении. Помимо ДНК, он взамодействует и с белковой цепью из соседней ячейки, образуя своеобразный димер. Таким образом, при анализа pdb-структуры нельзя забывать, что исходно мы смотрим только на малую часть кристалла и что существует еще множество ячеек, каждая из которых влияет на расположение молекул в соседних и поддерживает структуру кристалла.
Белок 1QD9
Кристаллографческие характеристики структуры можно найти в поле CRYST1 в pdb-файле. Оттуда можно узнать:
- размер ячейки (длины направляющих векторов): 53.350, 53.350, 204.960
- углы между векторами: 90.00, 90.00, 120.00
- число молекул в ячейке: 18
- кристаллографическая группа: P 65
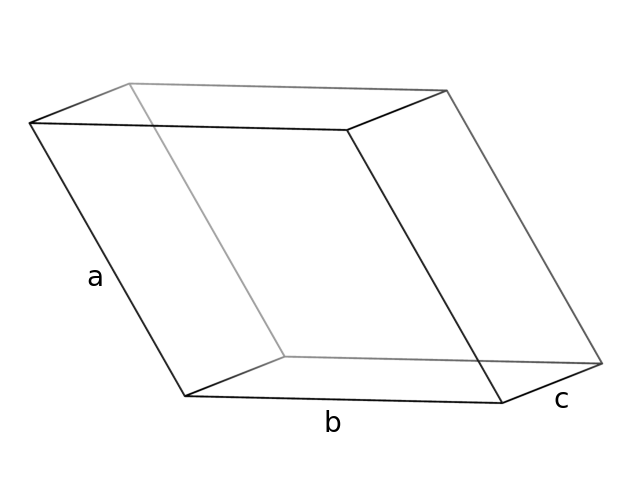
На картинке показана единичная кристаллографическая ячейка. "P" обозначает примитивную центровку ячейки (без дополнительных узлов). Эта ячейка имеет среднюю категорию (две стороны равны, два угла равны 90 градусов, а третий - 120), то есть у нее есть ось шестого порядка. Таким образом, симметрия кристалла гексагональная (образуется призма с шестиугольником в основании).
Рис.6: Кристаллографическая ячейка структуры 1QD9
Чтобы найти контакты белка с молекулами из соседних ячеек, с помощью команды symexp были восстановлены кристаллографические ячейки на расстоянии не более чем 4 А от исходной структуры. Таких ячеек оказалось десять.
Рис.7: Структура 1QD9 с востановленными соседними ячейками
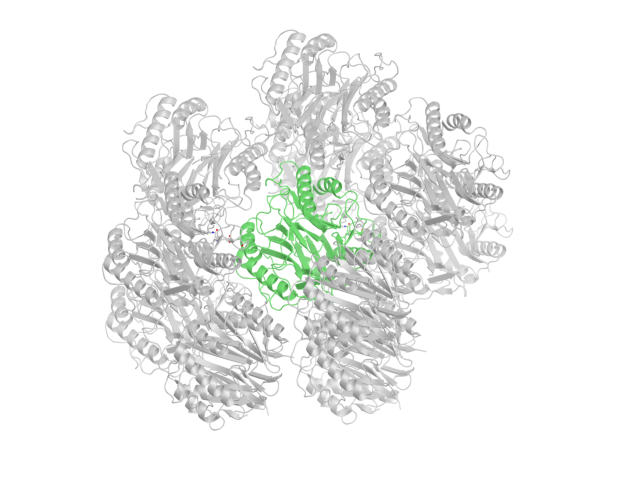
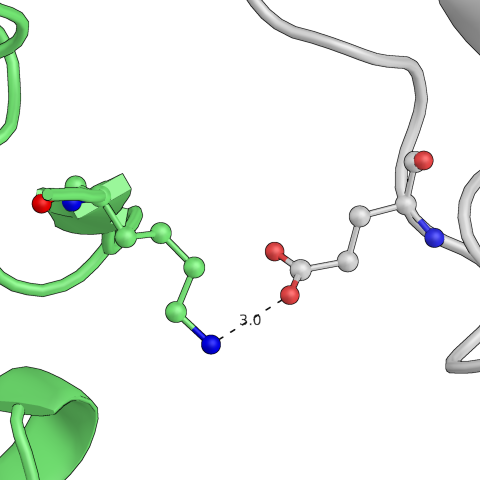
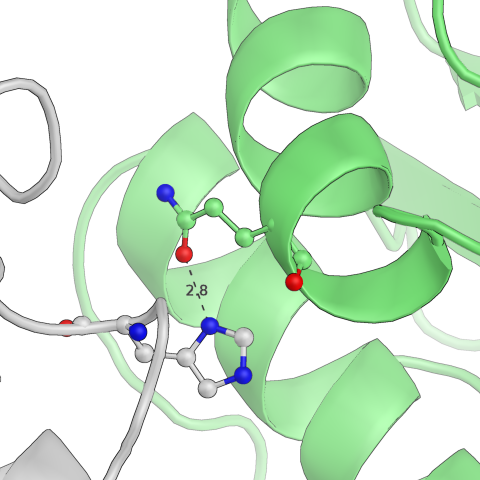
Рис.8: Примеры водородных связей в структуре
| Lys — Glu | Gln — His |
 |  |
Два белка из соседних кристаллических решеток скреплены целым рядом водородных связей и засчет этого выровнены друг относительно друга:
Рис.9: Примеры водородных связей в структуры
Glu - Asn, Asp - Asn, Glu - Lys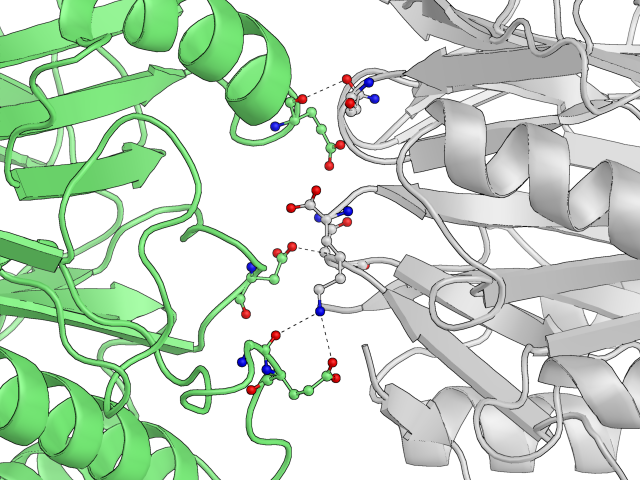
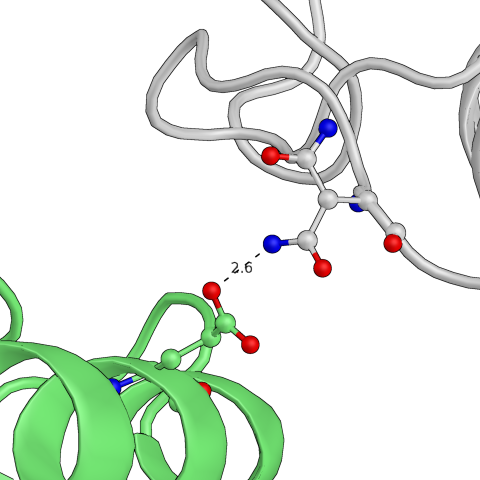
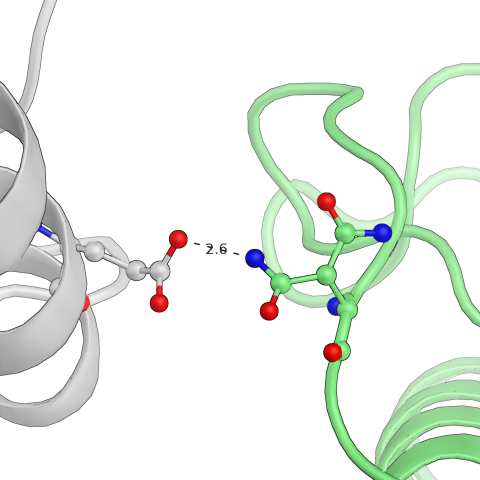
Естественно, встречаются аболютно "симметричные" контакты. Например, на рисунке ниже показан контакт, который представлен дважды. Первый раз взаимодействуют глутамат исходной "центральной" молекулы и аспарагин другой, а во второй раз - наоборот, но уже с другой стороны от центральной структуры. То же самое, как можно догадаться, есть во всех молекулах.
Рис.10: Пример "симметричного" водородного контакта в структуре
| Glu — Asn | Asn — Glu |
 |  |
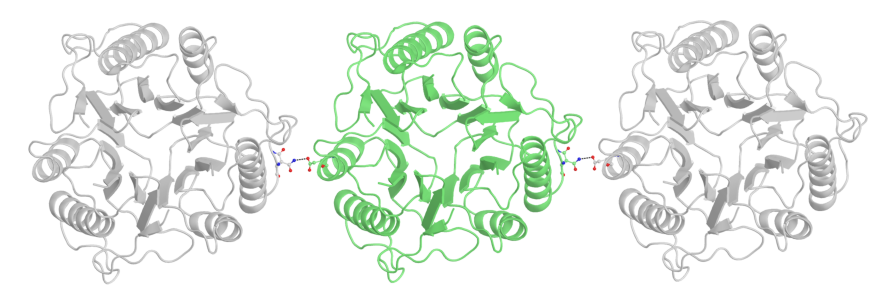
Ниже показан еще один пример такой симметрии. Она соответствует периодической структуре кристалла, ведь эти контакты повторяются между каждой парой ячеек.
Рис.11: Пример "симметричного" водородного контакта в структуре
 |
Выводы:
В структуре 1QD9 также присутствует множество межмолекулярных взаимодействий между белками из разных кристаллографических ячеек. Эти взаимодействия имеют периодический характер, повторяясь для каждой ячейки, что соответствует периодическому строению кристалла. Влияние межмолекулярных контактов на конформацию структуры и её ориентацию в пространстве является естественным следствием кристаллического строения образца для рентгеноструктурного анализа, и его нельзя упускать при анализе структур.