Практикум 3.
Задание 1. Предсказание вторичной структуры тРНК 1i9v
Предсказание структуры тРНК с помощью find_pair уже проводилось в предыдущем практикуме. В этот раз задание состояло в сравнении разных способов предсказания вторичной структуры тРНК - find_pair (поиск всех канонических и возможных неканонических пар), einverted (поиск инвертированных повторов) и RNAFold (алгоритм Зукера). Видно, что поиск инвертированных повторов достаточно плохо работает даже при подборе параметров, а find_pair и RNAFold выдают практически идентичные результаты уже при стандартных настройках RNAFold (первое по счёту предсказание). При этом RNAFold в моём случае лучше определил антикодоновый стебель, чем find_pair.

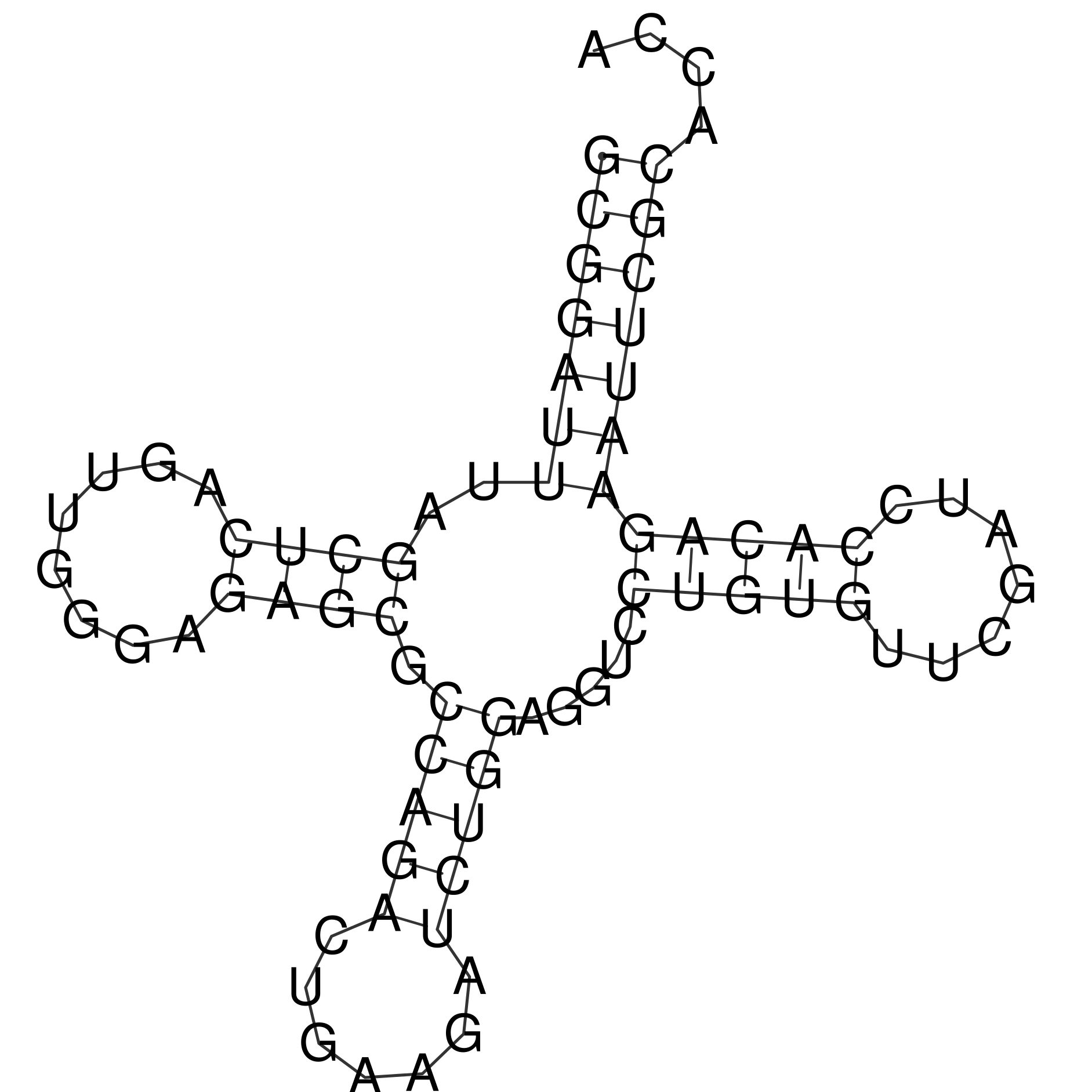
Результаты сравнения приведены в таблице 1.
| Участок структуры | Позиции в структуре (по результатам find_pair) |
Результаты предсказания с помощью einverted |
Результаты предсказания по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 5'-01-07-3' 5'-66-72-3' Всего 7 пар |
5'-05-07-3' 5'-66-68-3' Всего 3 пары |
7 пар |
| D-стебель | 5'-10-13-3' 5'-22-25-3' Всего 4 пары |
0 | 4 пары |
| T-стебель | 5'-49-53-3' 5'-61-65-3' Всего 5 пар |
5 пар | 5 пар |
| Антикодоновый стебель | 5'-27-30-3' 5'-40-43-3' Всего 4 пары |
0 | 5'-27-31-3' 5'-39-43-3' Всего 5 пар |
| Общее число канонических пар нуклеотидов | 21 | 8 | 22 |
Задание 2. Поиск ДНК-белковых контактов в структуре 1mdm (цепь белка B)
Упражнение 1
Упражнение 2
Скрипт для определения контактов.
Видно, что остатки дезоксирибозы в основном образуют неполярные контакты, что вполне ожидаемо. Однако фосфаты, если пользоваться предложенными критериями, также образуют в основном неполярные контакты, что достаточно странно - кислород, окружающий фосфор, обычно препятствует образованию неполярных контактов. С основаниями со стороны малой бороздки контактов не было найдено вовсе. В целом с азотистыми основаниями меньше контактов, чем с любым из компонентов сахарофосфатного остова, что вполне логично.
| Контакты атомов белка с | Полярные | Неполярные | Всего |
|---|---|---|---|
| остатками 2'-дезоксирибозы | 3 | 13 | 16 |
| остатками фосфорной кислоты | 8 | 18 | 26 |
| остатками азотистых оснований со стороны большой бороздки |
4 | 1 | 5 |
| остатками азотистых оснований со стороны малой бороздки |
0 | 0 | 0 |
Упражнение 3
Мы рассматриваем только белковую цепь B (цепь белка указана в скобках). Видно, что контактов с ДНК всего 10.
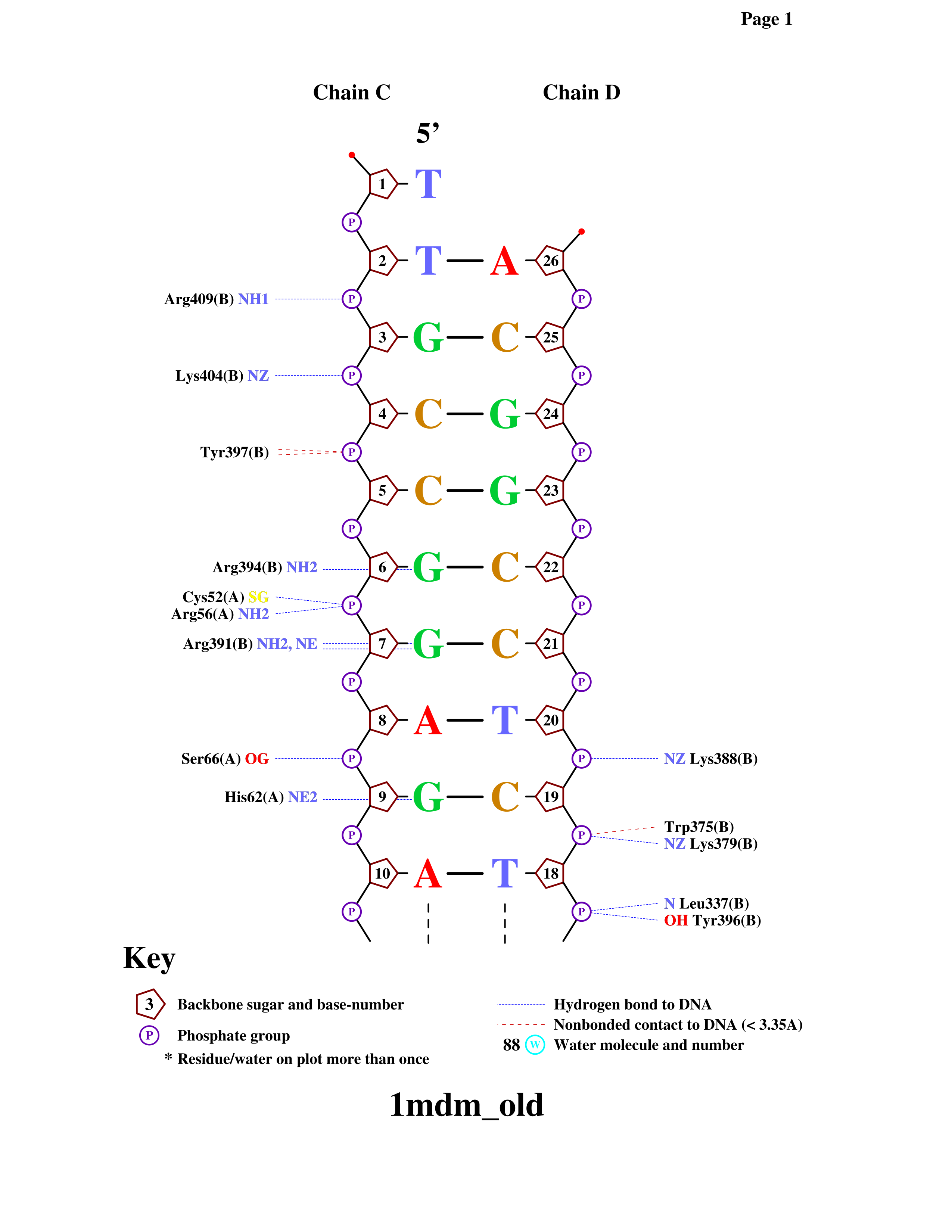
Упражнение 4
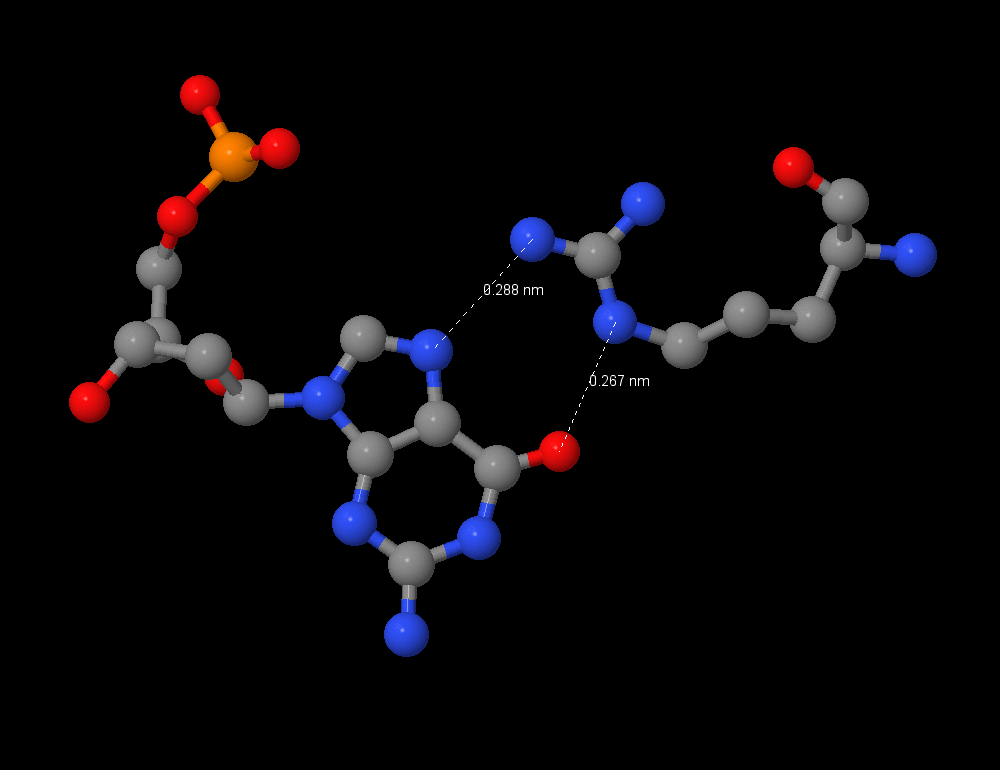
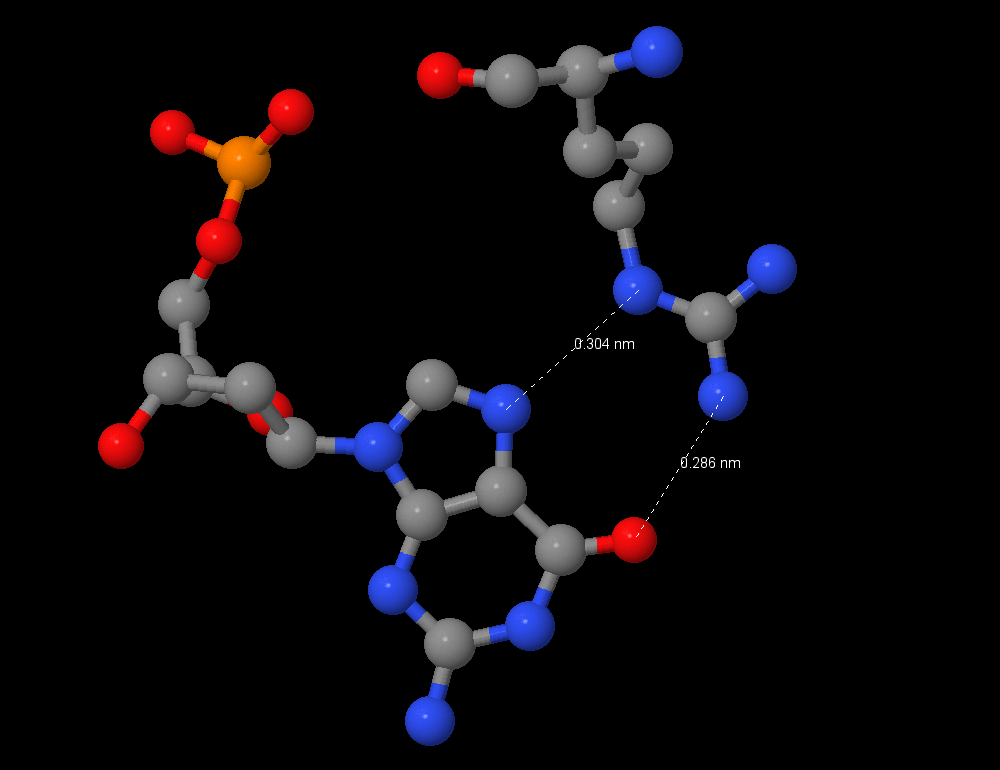
Из nucplot видно, что больше всего контактов с ДНК у Arg 391:B с G7:C. Это специфическое взаимодействие аргинина с гуанином, достаточно известное - с пиримидином аргинин не будет взаимодействовать из-за размера, а с аденином - из-за NH2-группы (донор водорода) вместо O (акцептор водорода). Однако есть и другой аргинин Arg 394:B, взаимодействующий с G6:C. Nucplot показывает только одну связь, однако я считаю, что это также специфическое взаимодействие аргинин-гуанин, что можно видеть, сравнив рисунки 4 и 5. Возможно, при расстоянии более 3 ангстрем взаимодействие не распознаётся nucplot. Таким образом, оба этих остатка важны для распознавания ДНК.