Определение вторичной структуры

|

|

|

|

|

|

|

|

|
Определение вторичной структуры с помощью DSSP и STRIDE
Для работы с этим заданием использовалась структура 2yjp ABC-транспортерного периплазматического связывающего аминокислоты белка. Данный белок содержит в своей структуре как α-спирали, так и β-листы.
В структуре файла PDB предусмотрена аннотация вторичной структуры. Вот как она выгляит в файле 2yjp.pdb:
HELIX 2 2 GLY A 72 GLY A 87 1 16
HELIX 3 3 SER A 88 ASP A 90 5 3
HELIX 4 4 GLU A 98 ALA A 100 5 3
HELIX 5 5 ASN A 101 SER A 108 1 8
HELIX 6 6 THR A 121 GLU A 126 1 6
HELIX 7 7 ASP A 151 LYS A 156 5 6
HELIX 8 8 THR A 166 HIS A 176 1 11
HELIX 9 9 GLN A 186 ASP A 196 1 11
HELIX 10 10 ASN A 206 ASN A 216 1 11
HELIX 11 11 ASN A 240 GLY A 258 1 19
HELIX 12 12 GLY A 258 LEU A 268 1 11
HELIX 13 13 LEU A 268 GLY A 273 1 6
HELIX 14 14 GLU A 274 VAL A 276 5 3
HELIX 15 15 LYS A 277 LEU A 282 1 6
HELIX 16 16 THR B 40 GLY B 48 1 9
HELIX 17 17 GLY B 72 GLY B 87 1 16
HELIX 18 18 SER B 88 ASP B 90 5 3
HELIX 19 19 GLU B 98 ALA B 100 5 3
HELIX 20 20 ASN B 101 SER B 108 1 8
HELIX 21 21 GLU B 123 GLU B 126 5 4
HELIX 22 22 ASP B 151 LEU B 155 5 5
HELIX 23 23 THR B 166 HIS B 176 1 11
HELIX 24 24 GLN B 186 ASP B 196 1 11
HELIX 25 25 ASN B 206 ASN B 216 1 11
HELIX 26 26 ASN B 240 GLY B 258 1 19
HELIX 27 27 GLY B 258 LEU B 268 1 11
HELIX 28 28 LEU B 268 GLY B 273 1 6
HELIX 29 29 GLU B 274 VAL B 276 5 3
HELIX 30 30 LYS B 277 LEU B 282 1 6
HELIX 31 31 THR C 40 GLY C 48 1 9
HELIX 32 32 GLY C 72 GLY C 87 1 16
HELIX 33 33 GLU C 98 ALA C 100 5 3
HELIX 34 34 ASN C 101 SER C 108 1 8
HELIX 35 35 THR C 121 GLU C 126 1 6
HELIX 36 36 ASP C 151 LYS C 156 5 6
HELIX 37 37 THR C 166 HIS C 176 1 11
HELIX 38 38 GLN C 186 ASP C 196 1 11
HELIX 39 39 ASN C 206 GLU C 215 1 10
HELIX 40 40 ASN C 240 GLY C 258 1 19
HELIX 41 41 GLY C 258 LEU C 268 1 11
HELIX 42 42 LEU C 268 GLY C 273 1 6
HELIX 43 43 GLU C 274 VAL C 276 5 3
HELIX 44 44 LYS C 277 LEU C 282 1 6
SHEET 1 AA 3 VAL A 92 LEU A 96 0
SHEET 2 AA 3 ILE A 50 VAL A 54 1 O ILE A 50 N GLU A 93
SHEET 3 AA 3 LEU A 113 ILE A 114 1 O LEU A 113 N GLY A 53
SHEET 1 AB 3 ASP A 57 LYS A 58 0
SHEET 2 AB 3 GLY A 62 VAL A 64 -1 O GLY A 62 N LYS A 58
SHEET 3 AB 3 ASN A 70 GLN A 71 -1 O GLN A 71 N TYR A 63
SHEET 1 AC 2 VAL A 128 PHE A 130 0
SHEET 2 AC 2 ALA A 235 GLN A 237 -1 O VAL A 236 N ASP A 129
SHEET 1 AD 2 MET A 135 VAL A 137 0
SHEET 2 AD 2 GLU A 230 ILE A 232 -1 O GLU A 230 N VAL A 137
SHEET 1 AE 5 LYS A 180 PHE A 184 0
SHEET 2 AE 5 THR A 159 ASN A 163 1 O LEU A 160 N LEU A 182
SHEET 3 AE 5 ALA A 201 ASP A 205 1 O ALA A 201 N LEU A 161
SHEET 4 AE 5 LEU A 139 PRO A 144 -1 O GLY A 140 N HIS A 204
SHEET 5 AE 5 PHE A 219 ILE A 223 -1 O GLU A 220 N SER A 143
SHEET 1 BA 3 VAL B 92 LEU B 96 0
SHEET 2 BA 3 ILE B 50 VAL B 54 1 O ILE B 50 N GLU B 93
SHEET 3 BA 3 LEU B 113 ILE B 114 1 O LEU B 113 N GLY B 53
SHEET 1 BB 3 ASP B 57 LYS B 58 0
SHEET 2 BB 3 GLY B 62 VAL B 64 -1 O GLY B 62 N LYS B 58
SHEET 3 BB 3 ASN B 70 GLN B 71 -1 O GLN B 71 N TYR B 63
SHEET 1 BC 2 VAL B 128 PHE B 130 0
SHEET 2 BC 2 ALA B 235 GLN B 237 -1 O VAL B 236 N ASP B 129
SHEET 1 BD 2 MET B 135 VAL B 137 0
SHEET 2 BD 2 GLU B 230 ILE B 232 -1 O GLU B 230 N VAL B 137
SHEET 1 BE 5 LYS B 180 PHE B 184 0
SHEET 2 BE 5 THR B 159 ASN B 163 1 O LEU B 160 N LEU B 182
SHEET 3 BE 5 ALA B 201 ASP B 205 1 O ALA B 201 N LEU B 161
SHEET 4 BE 5 LEU B 139 PRO B 144 -1 O GLY B 140 N HIS B 204
SHEET 5 BE 5 PHE B 219 ILE B 223 -1 O GLU B 220 N SER B 143
SHEET 1 CA 3 VAL C 92 LEU C 96 0
SHEET 2 CA 3 ILE C 50 VAL C 54 1 O ILE C 50 N GLU C 93
SHEET 3 CA 3 LEU C 113 ILE C 114 1 O LEU C 113 N GLY C 53
SHEET 1 CB 3 ASP C 57 LYS C 58 0
SHEET 2 CB 3 GLY C 62 VAL C 64 -1 O GLY C 62 N LYS C 58
SHEET 3 CB 3 ASN C 70 GLN C 71 -1 O GLN C 71 N TYR C 63
SHEET 1 CC 2 VAL C 128 PHE C 130 0
SHEET 2 CC 2 ALA C 235 GLN C 237 -1 O VAL C 236 N ASP C 129
SHEET 1 CD 5 LYS C 180 PHE C 184 0
SHEET 2 CD 5 THR C 159 ASN C 163 1 O LEU C 160 N LEU C 182
SHEET 3 CD 5 ALA C 201 ASP C 205 1 O ALA C 201 N LEU C 161
SHEET 4 CD 5 MET C 135 PRO C 144 -1 O GLY C 140 N HIS C 204
SHEET 5 CD 5 PHE C 219 ILE C 232 -1 O GLU C 220 N SER C 143
CISPEP 1 ALA A 38 ALA A 39 0 -12.97
CISPEP 2 PRO A 59 PRO A 60 0 7.26
CISPEP 3 PRO B 59 PRO B 60 0 1.05
CISPEP 4 PRO C 59 PRO C 60 0 7.07
На рисунках 1 и 2 показано cartoon-представление белка 2yjp с вторичными структурами, определенными в PDB и предсказанными с помощью dssp, встроенного в PyMol. Видно, что в целом элементы структуры, определенные dssp, меньше, чем аннотированные в PDB, особенно это касается бета-листов.

Рис.1. Элементы вторичной структуры ABC-транспортерного периплазматического связывающего аминокислоты белка, указанные в аннотации pdb. Петли показаны зеленым, α-спирали показаны красным, β-листы показаны желтым.
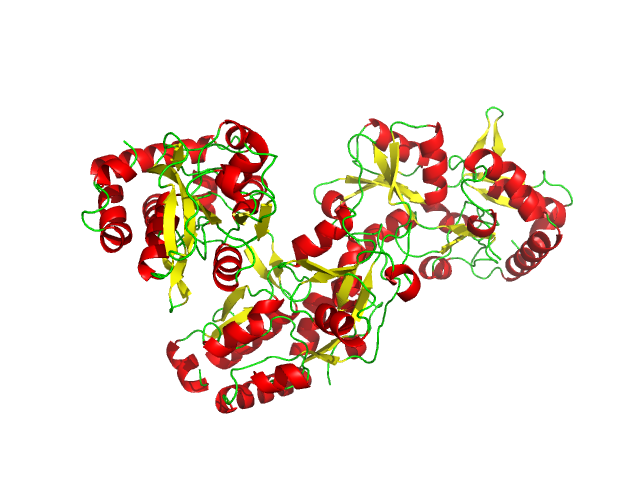
Рис.2. Элементы вторичной структуры ABC-транспортерного периплазматического связывающего аминокислоты белка, предсказанные dssp. Петли показаны зеленым, α-спирали показаны красным, β-листы показаны желтым.
Далее для удобства будем рассматривать только цепь белка А.
С помощью веб-сервиса STRIDE была построена карта вторичной структуры для белка 2yjp. На этой карте альфа-спирали показаны красным, 3-10 спирали синим, бета-листы зеленым. Выдача STRIDE для 2yjp доступна по ссылке. Некоторые элементы, предсказанные STRIDE, были сравнены с соответствующими элементами из аннотации в файле PDB. Результаты представлены в таблице 1.
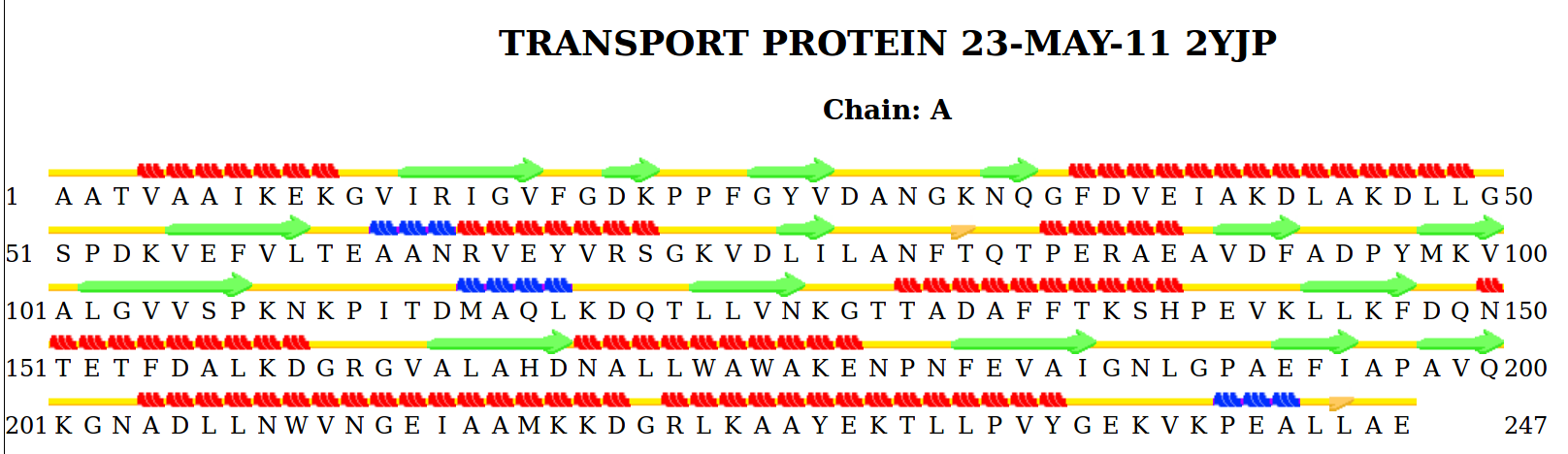
Рис.3. Элементы вторичной структуры ABC-транспортерного периплазматического связывающего аминокислоты белка, предсказанные STRIDE. Петли показаны желтым, α-спирали показаны красным, β-листы показаны зеленым, спирали 3-10 показаны синим, участки β-мостов показаны оранжевым.
| Тип вторичной структуры | Начало в pdb | Начало в STRIDE | Конец в pdb | Конец в STRIDE | Комментарий |
| α-спираль | 72E | 73F | 87G | 86L | Stride более аккуратен в своих предположениях |
| α-спираль | 101N | 102R | 108S | 108S | Stride более аккуратен в своих предположениях |
| β-лист | 139L | 139L | 144P | 144P | Stride предсказал правильно |
| β-лист | 219F | 219F | 223I | 223I | Stride предсказал правильно |
| Спираль 3-10 | 98E | 99A | 100A | 101N | Аннотация в PDB сдвинута на 1 остаток назад. В PDB HELIX не подразделяется альфа и 3-10 спираль, но сам момент перехода отслежен, пускай и со сдвигом на один остаток относительно предсказания Stride |
| Цис-конформация | 59P | - | 60P | - | Stride не находит участки в цис-конформации |
| β-мост | - | 119T | - | 119T | В PDB нет аннотации β-моста |
Сопоставление выдачи SheeP с белком
Для построения карты β-листов белка использовался сервис SheeP. Схема предсказанного листа и изображение представлены на рис. 4 и рис. 5.

Рис.4. Схема предсказанного β-листа.
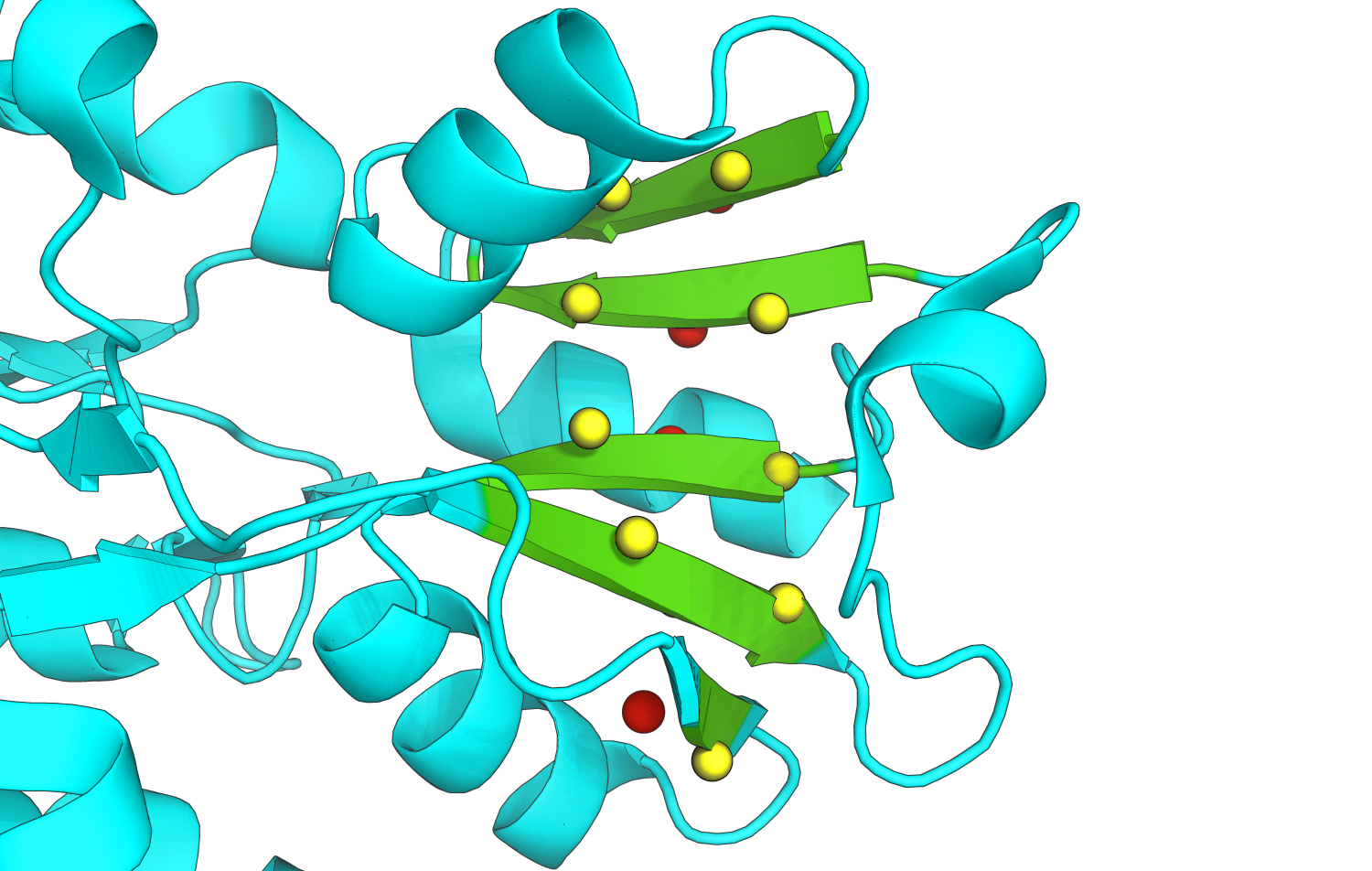
Рис.5. Изображение выбранного листа в структуре 2yjp. Остатки, сходящие в состав листе, покрашены в зеленый. Для трех хребтов показано положение соответствующих С-α атомов с сохранением цветовой кодировки из SheeP.
Перечислим остатки, включенные три центральных хребта данного листа. Левый желтый хребет: Lys183-Val162-Ala203-Val141. Центральный розовый хребет: Leu182-Leu161-Leu202-Val142-Val221. Правый желтый хребет: Leu181-Leu160-Ala201-Ser143-Glu220.
По цвету, в который SheeP покрасил хребты, можно определить, в какую сторону они смотрят — в сторону гидрофобного ядра или к поверхности. Для определения, красные или желтые хребты повернуты к гидрофобному ядру, проведем оценку аминокислотного состава. 6 из 11 остатков красных хребтов гидрофобны, в то время как в желтом — 7 из 9, что больше. Значит, к гидрофобному ядру повернуты желтые хребты. Как видно из рисунка 4, правый красный хребет обращен к растворителю, т.е. гидрофилен.
С помощью консольной версии STRIDE на kodomo был получен список водородных связей для участка, соответствующего карте выше. Полученная выдача приведена по ссылке. Аннотация водородных связей с помощью консольного STRIDE совпала с аннотацией встроенными инструментами PyMol. Водородные связи, определенные PyMol, показаны на рис.6.
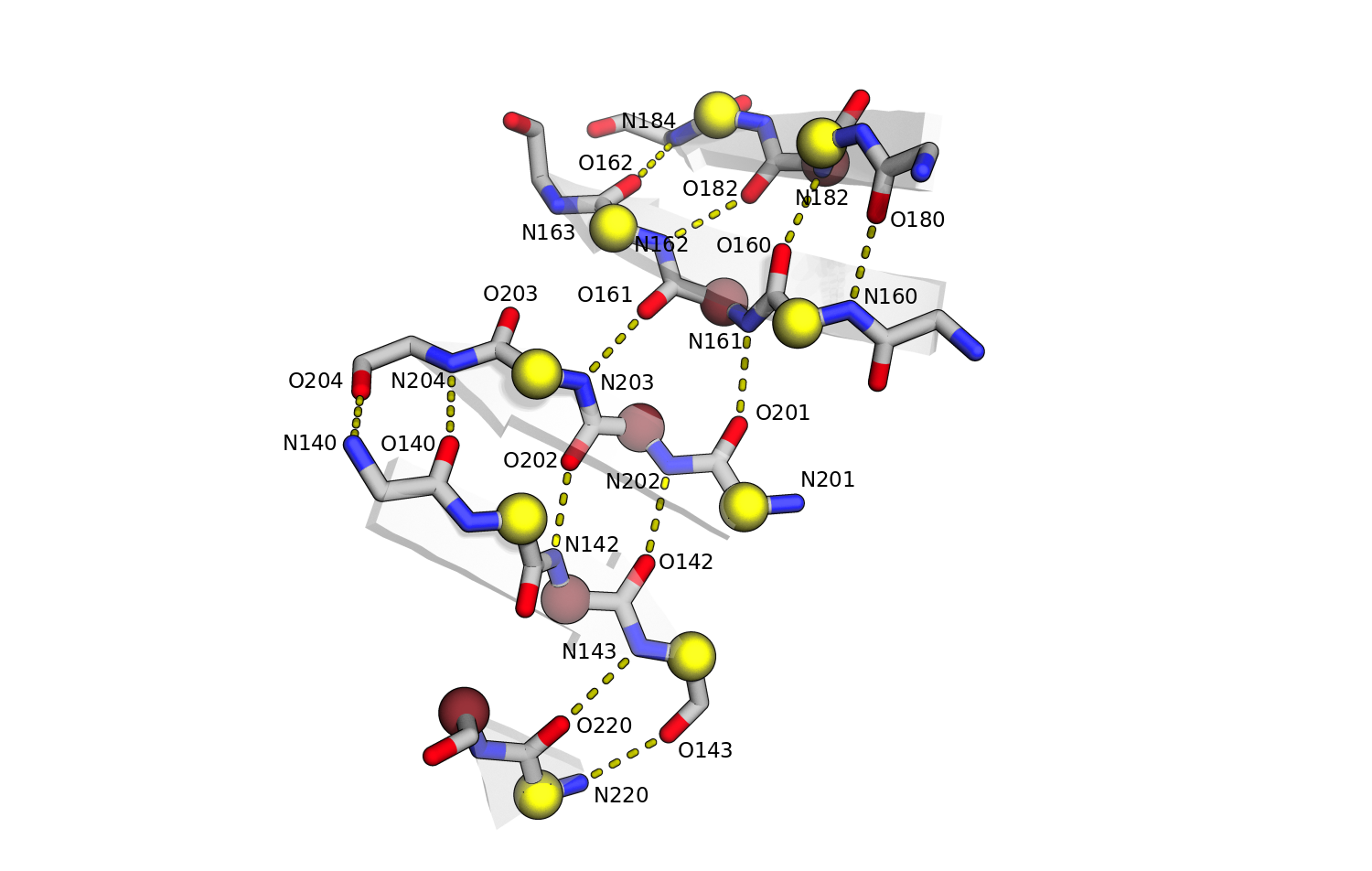
Рис.6. Водородные связи в β-листе. С-α атомы выделены шариками и покрашены по выдаче STRIDE. Водородные связи показаны пунктирными линиями.
Как видно из рис.6, явных нерегулярностей в структуре нет, за исключением отсутствия связи N163-O203.
Последнее изменение: 24.09.2014