 8 (926) 907 94 08
8 (926) 907 94 08
Всё на свете является чудом!
Алгоритмы реконструкции деревьев
Ранее в программе
JalView, используя метод Neighbour Joining, мы построили 2 неукорененных
филогенетических дерева. Теперь укореним одно из деревьев в среднюю
точку, но для интереса сделаем это для обоих деревьев ("NJ
Using % Identity" и "NJ Using
BLOSUM62").
Делать это с деревьями, построенными методом UPGMA
(Average Distance в программе JalView), не имеет
смысла, так как в отличие от метода NJ, они
уже укорененные и неметрические (с топологией, но без длин ветвей).
Итак, чтобы укоренить в среднюю точку дерево,
построенное методом Neighbour Joining (NJ), воспользуемся программой retree пакета PHYLIP.
О том как это сделать достаточно подробно рассказано в самом задании:
"Скопируйте
файл retree.exe с диска P в свою рабочую директорию. Скопируйте файл с
деревом в Newick-формате в файл с именем intree (именно так, без
расширения!) Запустите retree (из командной строки Windows; например
можно в Far manager навести курсор на файл retree.exe и нажать
В дополнение, отметим, что, нажав "
После чего, нажали на "M", чтобы укоренить построенное дерево в центральную точку (ниже приведена строчка из справки, показывающая, что нам нужно было нажать именно на "M")

Далее представлены и рассмотрены изображения соответствующих укорененных деревьев, полученные программой MEGA:
1) Neighbour Joining Using % Identity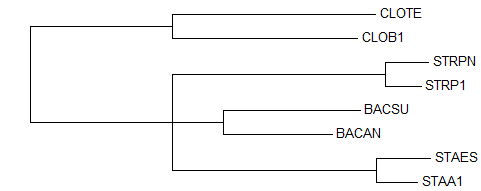 → дерево до укоренения
→ дерево до укоренения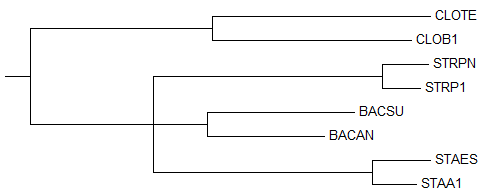 → укорененное дерево
→ укорененное дерево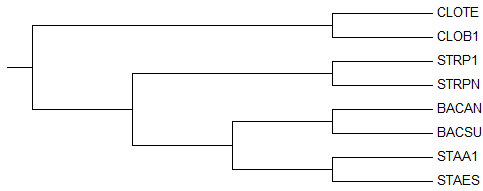 →
правильное дерево
→
правильное деревоУкоренение произошло правильно, в ветвь {CLOTE, CLOB1} против {STRPN, STRP1, BACSU, BACAN, STAES, STAA1}, несмотря на то, что изначально дерево, построенное методом
"NJ Using % Identity" не совпало с правильным из-за отсутствия соединения ветвей {BACAN, BACSU} и {STAA1, STAES} друг с другом.Деревья до и после укоренения отличаются только длиной ветви {CLOTE, CLOB1}. 2) Neighbour Joining Using BLOSUM62
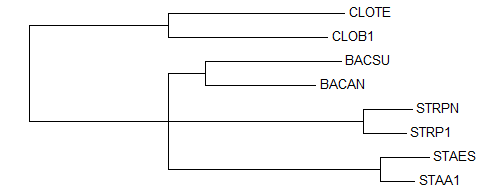 → дерево до укоренения
→ дерево до укоренения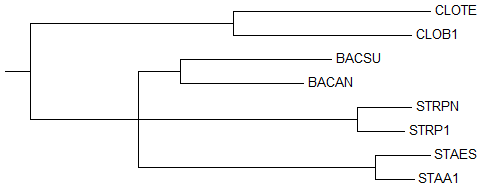 → укорененное дерево →
правильное дерево
→ укорененное дерево →
правильное деревоДерево до укоренения, построенное этим методом (
"NJ Using BLOSUM62"), совпадает с деревом до укоренения, построенным предыдущим методом ("NJ Using % Identity"), независимо от того, что ветви {BACSU, BACAN} и {STRPN, STRP1} поменялись местами, это ничего не меняет.Укоренение, как и в предыдущем случае произошло правильно, в ту же ветвь {CLOTE, CLOB1} против {STRPN, STRP1, BACSU, BACAN, STAES, STAA1}. Тут деревья до и после укоренения отличаются тоже только длиной ветви {CLOTE, CLOB1}. 2. Использование внешней группы
Деревья, построенные методом максимальной экономии ("Maximum parsimony")
невозможно укоренить в среднюю точку, так как метод не реконструирует
длины ветвей, поэтому самый длинный путь от листа к листу не посчитать,
соответственно и среднюю точку никак не найти. Но можно воспользоваться
укоренением с помощью внешней группы.
В качестве внешней группы выбираем белок того же семейства (EFG)
из кишечной палочки (efg_ecoli). Для
этого к файлу с
невыровненными последовательностями
белков фирмикут с функцией "Фактор элонгации трансляции G"
добавляем последовательность белка из кишечной палочки, используя
команду cat:
Затем выравниваем полученные последовательности с помощью программы muscle:
muscle -in all_bacteria_and_ecoli.fasta -out alignment_all_bacteria_and_ecoli.fasta
Результат выравнивания импоруем в программу MEGA, предварительно отредактировав имена последовательностей (оставив только мнемонику видов).
Теперь строим дерево (Maximum Parsimony в меню Phylogeny) и укореняем его в ветвь, ведущую к ECOLI (Subtree > Root). Используя кнопку Show Subtree Separately получаем изображение дерева без ECOLI. Полученное изображение укорененного дерева приведено ниже:
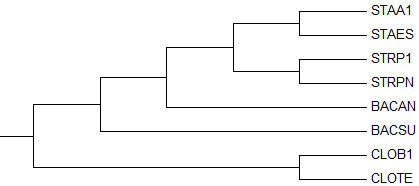 → укорененное дерево →
правильное дерево
→ укорененное дерево →
правильное деревоВ очередной раз укоренение успешно прошло в ветвь {CLOTE, CLOB1}
против {STRPN, STRP1, BACSU, BACAN, STAES, STAA1}.
Но реконструирование дерева методом максимальной экономии с
использованием внешней группы произошло с ошибками - ветвь
Бутстрэп-анализ филогении отобранных белков проведем, используя один
из методов, доступных в программе MEGA.
Например,
ниже приведены изображения деревьев, полученные в результате бутстрэп-анализа белков с использованием метода UPGMA (число реплик было
установлено равным 100).
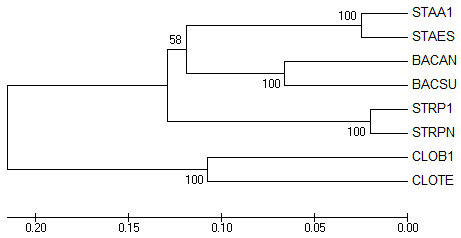 → original tree
→ original tree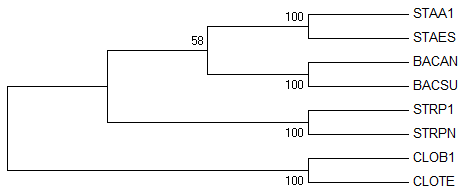 → bootstrap consensus tree →
правильное дерево
→ bootstrap consensus tree →
правильное дерево
Оба полученных дерева (Original tree и Bootstrap consensus tree)
совпадают. Более того, эти деревья полностью совпадают и с правильным
деревом. Естественно все 5 нетривиальных ветвей у трех представленных
выше деревьев тоже одинаковые.
Неправильных ветвей нет, поэтому и сравнить поддержку мы не можем. Но
заметим, что у 4-х нетривиальных ветвей (все они конечно правильные)
максимальная поддержка (100%
бутстрэп-копий с данной ветвью).
| Главная | Об авторе | Учебные семестры | Проекты автора | Друзья | Ссылки партнеров | Extra | Контакты |
Mneff © 2011-2013