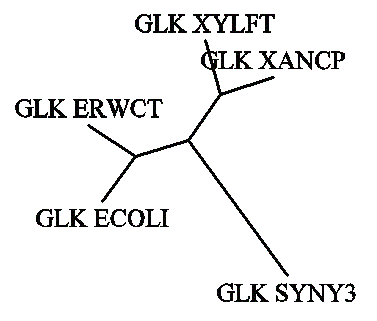
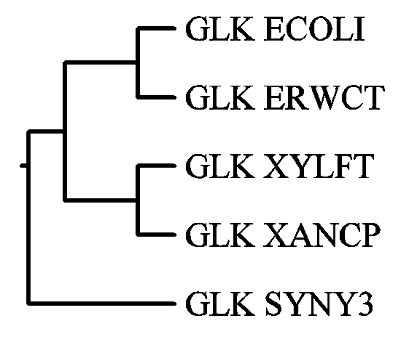
Филогенетические деревья, реконструированные разными способами
В работе были реконструированы филогенетические деревья по эталонному выравниванию доменов пяти различных белков глюкокиназы.
Эталонное выравнивание исследуемых аминокислотных последовательностей белков:
| * | 2 | 0 | * | 4 | 0 | * | 6 | 0 | * | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | L | K | _ | X | Y | L | F | T | _ | : | L | V | A | D | I | G | G | T | N | A | R | F | A | L | A | N | P | T | L | T | S | - | - | - | A | P | L | L | N | D | S | M | R | E | F | A | V | I | E | F | P | S | L | G | E | A | A | Q | H | Y | L | H | - | - | H | I | G | I | H | T | T | K | - | - | - | - | G | V | F | A | : | 6 | 1 | ||||
| G | L | K | _ | X | A | N | C | P | _ | : | L | V | A | D | I | G | G | T | N | A | R | F | A | L | A | D | I | D | A | S | V | - | - | - | - | P | L | L | D | D | T | C | R | E | F | A | V | V | E | F | G | S | L | G | E | A | A | R | Y | Y | L | D | - | - | Q | I | G | V | Q | A | T | K | - | - | - | - | G | V | F | A | : | 6 | 0 | ||||
| G | L | K | _ | E | R | W | C | T | _ | : | L | V | G | D | V | G | G | T | N | T | R | L | A | L | C | D | A | M | T | G | E | - | - | - | - | - | - | - | L | S | Q | I | E | T | Y | S | G | L | D | F | P | S | L | E | G | A | I | R | D | Y | L | D | S | R | Q | V | T | V | Q | D | - | - | - | - | - | - | A | C | I | A | : | 5 | 7 | ||||
| G | L | K | _ | E | C | O | L | I | _ | : | L | V | G | D | V | G | G | T | N | A | R | L | A | L | C | D | I | A | S | G | E | - | - | - | - | - | - | - | I | S | Q | A | K | T | Y | S | G | L | D | Y | P | S | L | E | A | V | I | R | V | Y | L | E | E | H | K | V | E | V | K | D | - | - | - | - | - | - | G | C | I | A | : | 5 | 7 | ||||
| G | L | K | _ | S | Y | N | Y | 3 | _ | : | L | A | G | D | I | G | G | T | K | T | I | L | A | L | V | T | I | N | E | S | S | P | G | L | A | R | P | V | T | L | F | E | Q | T | Y | S | S | P | A | F | P | D | L | V | P | M | V | Q | Q | F | R | Q | E | A | A | F | V | L | G | N | P | I | S | V | A | K | A | C | F | A | : | 7 | 0 | ||||
| L | D | G | G | T | A | L | L | A |
На основе матрицы попарных совпадений последовательностей в этом выравнивании была построена матрица эволюционных расстояний между исследуемыми последовательностями. Затем было построено филогенетическое дерево.
Выполнение работы отображено в Рабочей книге UPGMA.xls.
Правильная скобочная структура получившегося дерева:
((((GLK_ECOLI:0.12,GLK_ERWCT:0.12):0.31625, (GLK_XYLFT:0.12,GLK_XANCP:0.12):0.31625): 0.36125),GLK_SYNY3:0.36125);.
Была выполнена графическая визуализация дерева по его правильной скобочной структуре посредством программ drawtree и drawgram.
Первая из них строит неукоренённое дерево (соответствует левому рисунку), а вторая программа строит укоренённое дерево (соответственно, правому рисунку).
Полученные изображения:
|
|
Затем, были построены филогенетические деревья по методу ближайших соседей. Метод основан на множественном выравнивании аминокислотных последовательностей, выполненном посредством программы emma. Полученный в ходе работы файл NJ.dnd содержит скобочную структуру филогенетического дерева:
((GLK_XYLFT:0.11566,GLK_XANCP:0.11419):0.20104,(GLK_ERWCT:0.12383,GLK_ECOLI:0.11293):0.16846,GLK_SYNY3:0.39384);.
На её основе были построены неукоренённое и укоренённое деревья:
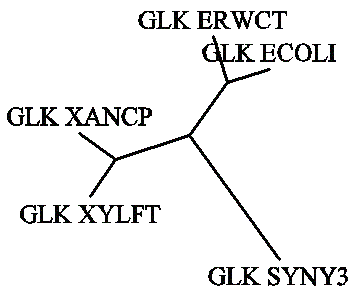 |
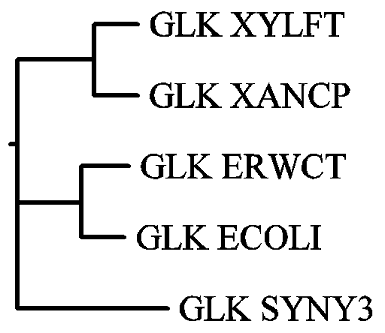 |
Сравнивая NJ-дерево с UPGMA-деревом, видно, что неукоренённые деревья практически совпадают, они отличаются лишь порядком расположения на плоскости рисунка двух ветвей филогенетического дерева, что несущественно и непринципиально. Укоренённые же деревья отличаются, т. к. в NJ-дереве исходный белок, в отличие от UPGMA-дерева, даёт начало не белку GLK_SYNY3 и одной ветви, которая затем разделится на 2 двойные ветви, а одной двойной ветви белков GLK_XYLFT и GLK_XANCP и ветви, делящейся на белок GLK_SYNY3 и ветвь белков GLK_ECOLI и GLK_ERWCT. При построении филогенетических деревьев разными способами получились неодинаковые сценарии эволюции белков.
На главную страницу второго семестра