Средства JMol для работы со структурами нуклеиновых кислот
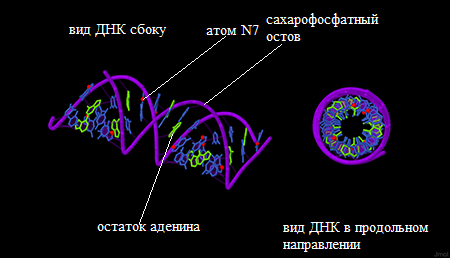
В упражнении 1 задания 2 нам было предложено овладеть инструментами работы со структурами ДНК, полученными в первом задании, в программе Jmol. Результат работы представлен на рис. 1.
Во втором упражнении задания 2 требовалось получить файлы PDB комплексов тРНК и ДНК с белками (мне достались структуры с идентификаторами 1exd и 1xbr соответственно). PDB-файлы можно скачать по следующим ссылкам:EXD, XBR
Модели были проверены на наличие разрывов
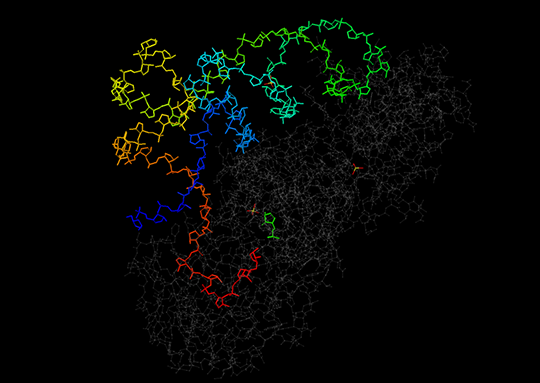
Разрыв в цепи тРНК структуры с идентификатором PDB 1exd был обнаружен только в режиме визуализации wireframe. Отдельный фрагмент тРНК состоит из одного нуклеотидного остатка. Расстояние между остатками фосфорной кислоты отдельного остатка и первого остатка на основной цепи составляет 1.005 нм, в то время как между первым остатком фосфорной кислоты и следующим через один 1.009 нм. Это может говорить о том, что величина разрыва составляет один нуклеотидный остаток.
С другой стороны состав азотистых оснований на 3’-конце основной цепи тРНК был соблюдён (последовательность CCA). По этой причине адениновый нуклеотид может быть остатком АТФ, используемой при активации аминокислот и дальнейшем их присоединении аминоацил-тРНК-синтетазой к различным. Структура тРНК изображена на рис. 2.
В структуре с идентификатором PDB 1xbr разрывов найдено не было. Структура представлена на рис. 3. Число нуклеотидных остатков в цепях ДНК совпадает (то есть, на концах цепей нет неспаренных оснований).
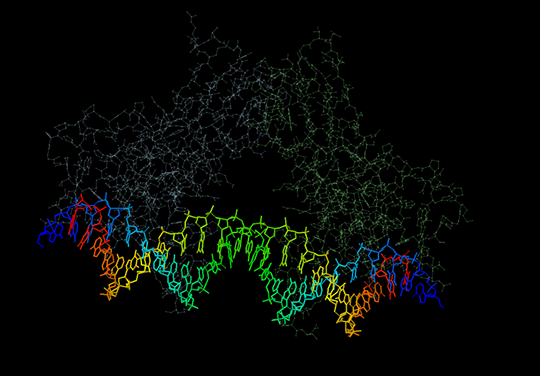
Также можно заметить, что цепи ДНК вдоль своего направления от 5’- к 3’-концу имеют одинаковый нуклеотидный состав. Это проиллюстрировано на рис. 4.
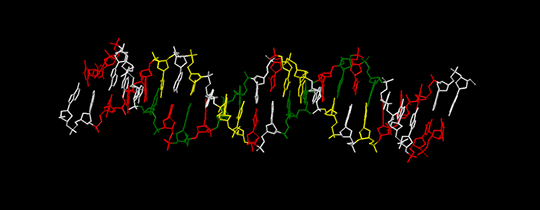
Сравнение структур 3-х форм ДНК с помощью средств JMol
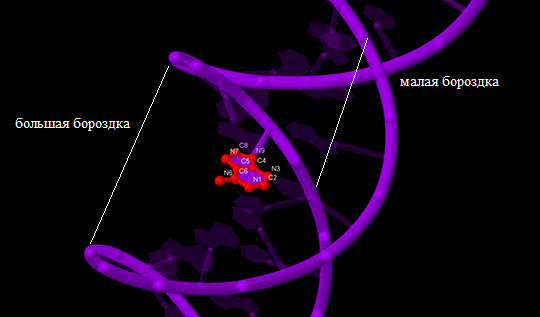
В упражнении 1 задания 3 нам было предложено найти большую и малую бороздки структур ДНК, полученных в задании 1, а также определить в какой из бороздок лежат те или иные атомы определённого азотистого основания (мне достался аденин).
Для анализа был выбран аденин остатка 34 цепи B. Результаты для В-формы ДНК представлены на рис. 5. В большой бороздке лежат атомы С6, N7, C5, N6. В малой бороздке лежат атомы N9, C4, N3, C2. Атомы N1 и C8 лежат на границе, разделяющей малую и большую бороздку.
В A-форме ДНК расположение атомов аденина остатка 34 цепи B меняется. При этом стоит отметить особенность большой бороздки данной формы ДНК: визуально она кажется уже, но в то же время имеет большую глубину, чем малая бороздка. В малой бороздке лежат атомы С6, N7, C5, N6. В большой бороздке лежат атомы N9, C4, N3, C2. При этом атомы N1 и C8 не относятся ни к одной из бороздок.
В Z-форме ДНК аденин отсутствует.
В упражнении 2 задания 3 было необходимо сравнить основные параметры спиралей ДНК различных форм. Результаты выполнения задания представлены в таблице 1.
| A-форма | B-форма | Z-форма | |
| Тип спирали (правая или левая) | правая | правая | левая |
| Шаг спирали (Å) | 28.03 | 33.75 | 43.5 |
| Число оснований на виток | 11 | 10 | 12 |
| Ширина большой бороздки (Å) | 9.63 | 20.58 | 9.97 |
| Ширина малой бороздки (Å) | 18.5 | 13.2 | 22.91 |
В упражнении 3 задания 2 требовалось измерить торсионные (двугранные) углы между атомами в одном нуклеотиде для А- и В-форм ДНК. Нам было предложено сравнить полученные результаты со значениями углов в презентации, любезно предоставленной Головиным А.В. Фрагмент презентации изображён на рис. 6.
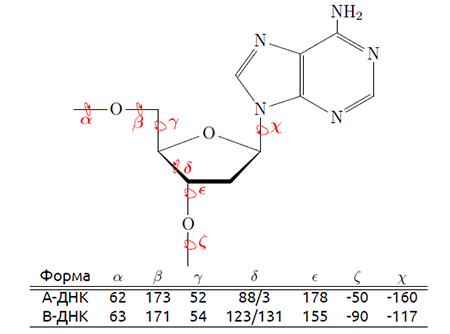
Результаты измерения углов представлены в таблице 2.
| Альфа | Бэтта | Гамма | Дэльта | Эпсилон | Пси | Кси | |
| А-форма | 64.1 | 174.8 | 41.7 | 79.1 | 100.3 | -75.1 | -157.2 |
| B-форма | 85.9 | 152.5 | 31.2 | 143.3 | 105.8 | -44.7 | -98.0 |
Были найдены средние значения торсионных углов для структур молекул ДНК и тРНК. Значения торсионных углов α, γ и δ 11 нуклеотидного остатка цепи 1 ДНК больше всего отличаются от своих средних значений (соответственно, 63° при среднем значении -28.1°, -121.4° при среднем значении 20.8°, 127.9° при среднем значении 138.8°).
В структуре тРНК не было обнаружено остатков с несколькими торсионными углами, чьи значения больше всего отличаются от среднего. Значения торсионного угла ε остатка 7 цепи 1 тРНК больше всего отличается от своего среднего значения (175.8° при среднем значении -123.2°).
В упражнении 2 задания 4 нам было предложено научиться определять структуру водородных связей. В файле .out, который был получен в упражнении 1, была найдена информация о водородных связях между азотистыми основаниями тРНК.
Strand I Strand II Helix 1 (0.010) B:.902_:[..G]G-----C[..C]:.971_:B (0.005) | 2 (0.012) B:.903_:[..G]G-----C[..C]:.970_:B (0.007) | 3 (0.011) B:.904_:[..G]G-----C[..C]:.969_:B (0.006) | 4 (0.012) B:.905_:[..G]G-----C[..C]:.968_:B (0.010) | 5 (0.010) B:.906_:[..U]U-----A[..A]:.967_:B (0.010) | 6 (0.013) B:.907_:[..A]Ax----U[..U]:.966_:B (0.004) | 7 (0.006) B:.949_:[..C]C-----G[..G]:.965_:B (0.011) | 8 (0.008) B:.950_:[..G]G-----C[..C]:.964_:B (0.004) | 9 (0.005) B:.951_:[..A]A-----U[..U]:.963_:B (0.004) | 10 (0.014) B:.952_:[..G]G-----C[..C]:.962_:B (0.005) | 11 (0.006) B:.953_:[..G]G----xC[..C]:.961_:B (0.005) | 12 (0.002) B:.954_:[..U]U-*--xA[..A]:.958_:B (0.008) | 13 (0.005) B:.955_:[..U]Ux**+xG[..G]:.918_:B (0.009) x 14 (0.007) B:.937_:[..A]A-*---U[..U]:.933_:B (0.003) | 15 (0.004) B:.938_:[..U]U-*---U[..U]:.932_:B (0.006) | 16 (0.006) B:.939_:[..U]U-----A[..A]:.931_:B (0.006) | 17 (0.005) B:.940_:[..C]C-*---G[..G]:.930_:B (0.012) | 18 (0.005) B:.941_:[..C]C-----G[..G]:.929_:B (0.007) | 19 (0.012) B:.942_:[..G]G-----C[..C]:.928_:B (0.010) | 20 (0.007) B:.943_:[..G]G-----C[..C]:.927_:B (0.003) | 21 (0.008) B:.944_:[..A]Ax*---A[..A]:.926_:B (0.015) | 22 (0.013) B:.910_:[..G]G-----C[..C]:.925_:B (0.006) | 23 (0.013) B:.911_:[..C]C-----G[..G]:.924_:B (0.006) | 24 (0.008) B:.912_:[..C]C----xG[..G]:.923_:B (0.007) | 25 (0.012) B:.913_:[..A]A-**+xG[..G]:.946_:B (0.008) | 26 (0.006) B:.914_:[..A]A-*--xA[..A]:.921_:B (0.008) | 27 (0.015) B:.915_:[..G]Gx**+xU[..U]:.948_:B (0.012) x 28 (0.029) B:.919_:[..G]G-----C[..C]:.956_:B (0.012) +
В составе тРНК присутствует 9 пар азотистых оснований с неканоническим типом взаимодействий. Они отмечены звёздочками (13 U-G, 15 U-U, 21 A-A, 25 A-G, 26 A-A, 27 G-U).
В поддержании третичной структуры тРНК участвуют остатки 919 и 956, 918 и 955, 913 и 946, 915 и 948.
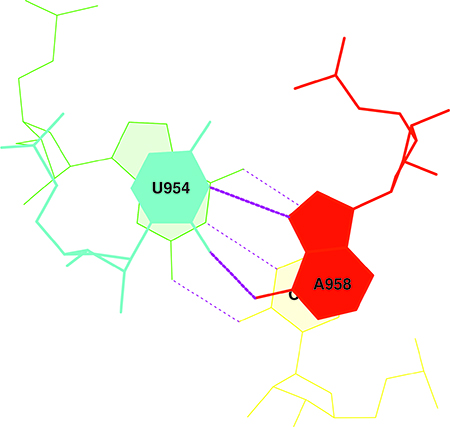
В упражнении 3 задания 4 было необходимо построить модель стекинг-взаимодействий между азотистыми основаниями с наибольшей площадью перекрывания.
Наибольшая площадь перекрывания наблюдается между парами оснований G-C 11 остатка и U-A 12 остатка. Изображение, иллюстрирующее стекинг-взаимодействия между остатками, можно увидеть на рис. 8.
Взаимная ориентация оснований была проверена с помощью Jmol (рис. 9).