| | | |
Оптимальное парное выравнивание. Алгоритмы |
Вернуться на страницу семестра Сравнение параметров глобального и локального выравниваний пары гомологичных белков семейства HSP70В данной работе мы сравниваем работу 2 алгоритмов выравнивания: в EMBOSS алгоритм глобального парного выравнивания Нидлмана — Вунша с аффинными штрафами за гэпы реализован в программе needle, локальное выравнивание по алгоритму Смита — Ватермана — в water. Были использованы белки археи Picrophilus torridus и эукариота C. elegans, уже рассматриваемые в предыдущей работе Предлагаю рассмотреть выравнивания, которые выдают оба алгоритма, сравнить эти выводы и заметить, как изменятся выводы алгоритмов при изменении штрафов за удлинение инделя при переходе от афинных штрафов к линейным. О том, как правильно понимать рисунки ниже: на каждой картинке приведён совмещенный вариант выравниваний по двум алгоритмам. 2 верхние последовательности получены алгоритмом Нидлмана — Вунша. 2 нижние (выделены красным пунктиром) алгоритмом Смита — Ватермана. Параметр консервативности в данных выравниваниях — 50% — для наглядности совпадения результатов разных алгоритмов. Для получения результата в формате fasta следует использовать запрос: needle(water) seqA.fasta seqB.fasta -auto -aformat fasta align.fasta Рис.1. Глобальное(needle) и локальное(water) выравнивание 1 при автоматических настройках (штраф за удлинение инделя = 0.5) Рис. 2. Глобальное и локальное выравнивание 2 (штраф = 5) Рис. 3. Глобальное и локальное выравнивание (штраф = 10)
Можно заметить, что при штрафах 0.5 и 5 выравнивания совпадают везде, кроме коцевого участка, начинающегося с 635 позиции; при штрафе, равном 10,
выравнивание менее полное и некоторые блоки, что выражены в первых двух случаях, отсутствуют.
Это наглядное доказательство преимущества алгоритмов с афинными штрафами перед линейными в случае гомологичных последовательностей.
Однако судить, какое из выравниваний (1 или 2) более правдоподобно, сложно.
Таблица 1. Параметры выравнивания 1 для разных алгоритмов
Таблица 2. Настройки выравнивания 1 (по умолчанию)
Команды для изменения параметров
Сравнение параметров локального выравнивания пары негомологичных белковТаблица 3. Выбранные негомологичные белки
Таблица 4. Локальное выравнивание гомологичных (1 столбец) и негомологичных (2 - 6 стоблец) белков
Комментарий к таблице: в локальном выравнивании гомологичных последовательностей процент абсолютно консервативных позиций вдвое больше, чем у негомологичных, функционально консервативных - много больше половины (почти 70%), процент гэпов наименьший, очень мало инделей с учётом длины выравнивания - 632. Рис. 4. Локальное(water) выравнивание гомологов Рис. 5. Локальное(water) выравнивание 1-2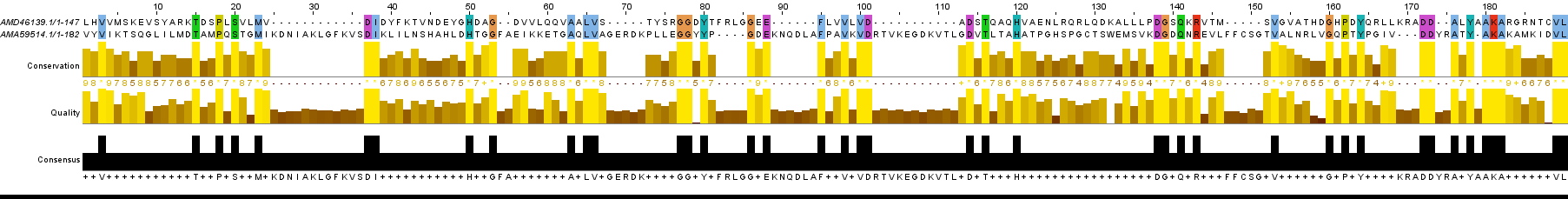 Рис. 6. Локальное(water) выравнивание 1-3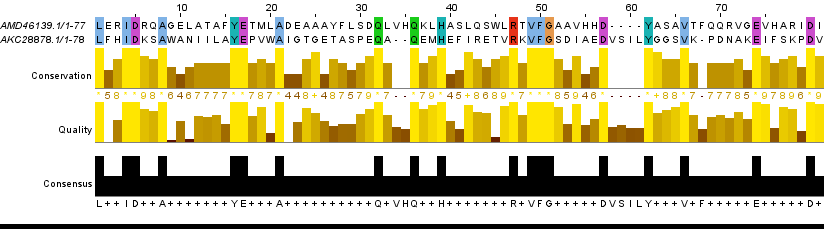 Рис. 7. Локальное(water) выравнивание 1-4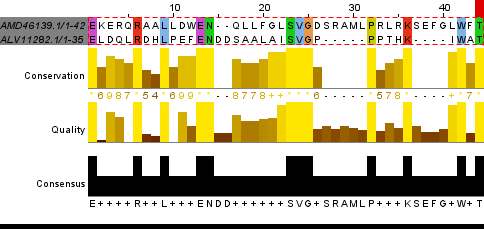 Рис. 8. Локальное(water) выравнивание 1-5 Рис. 9. Локальное(water) выравнивание 1-6 Отличия выравниваний, построенных разными программамиРис. 10. Выравнивания, построенные разными способами Пояснения к рисунку и комментарии: в выравнивании совмещены выравнивания, полученные 3 способами.
Первые 2 строки - парное выравнивание, полученное из множественного путем удаления всех последовательностей, кроме двух;
следующие 2 строки - глобальное выравнивание программой needle (EMBOSS), параметры по умолчанию.
Последние 2 строки - локальное выравнивание программой water (EMBOSS).
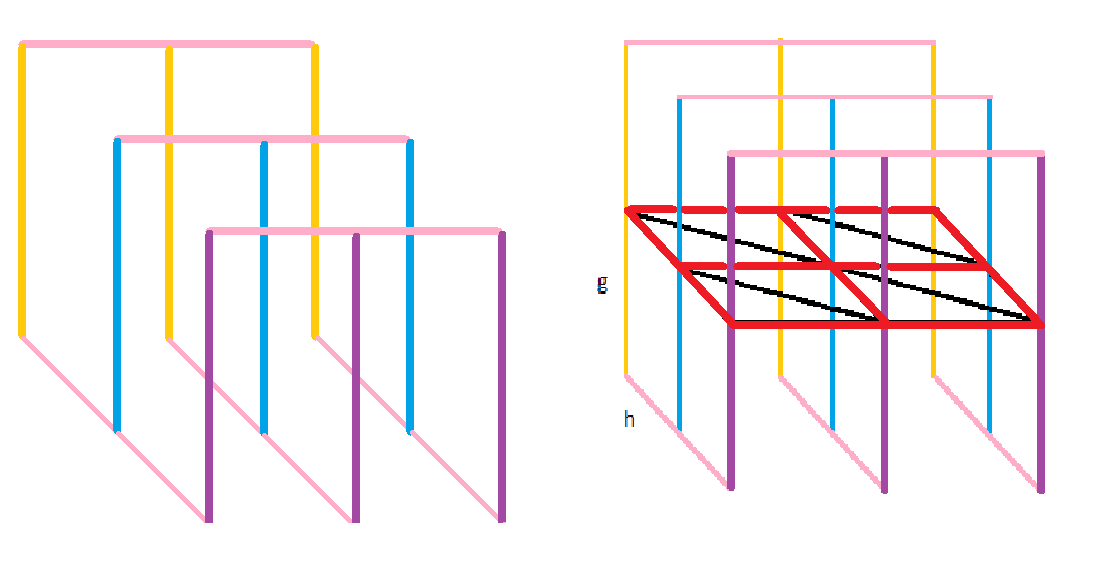
Ориентированный граф для построения глобального выравнивания с аффинными штрафами за инделиРис. 11. Граф глобального выравнивания с афинными штрафами. Собственная иллюстрация (участок выравнивания 2х2) и готовая [2].
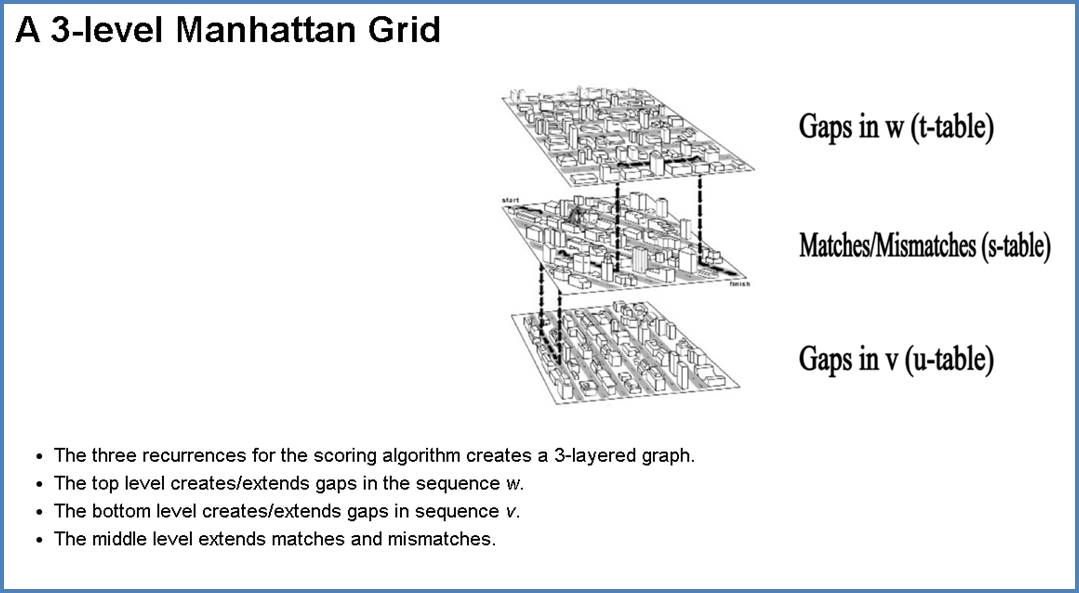
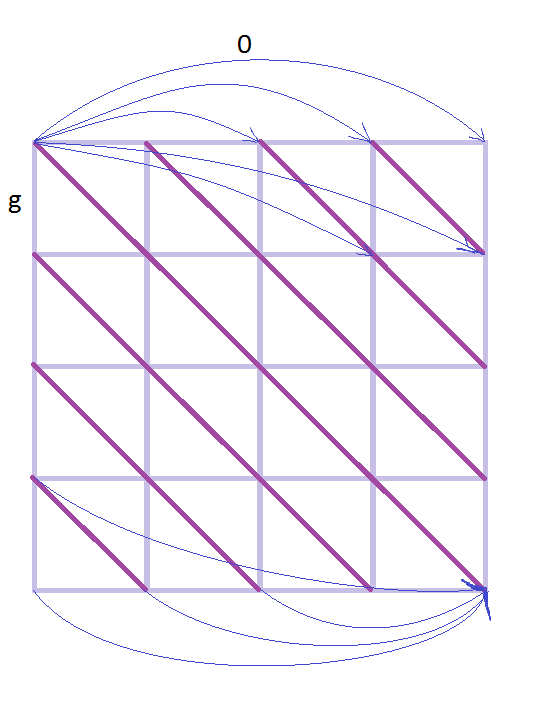
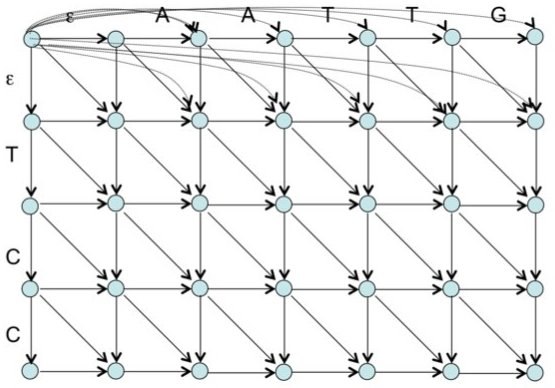
К известному графу для глобального выравнивания с линейным штрафом ("Манхэттен и Бродвеи") добавим 2 этажа: выше и ниже. Проведу аналогию с гипотетическими раем и адом для лучшего понимания: верхний этаж - рай, средний - бытие на земле, нижний - ад, штраф получают за состояние вне жизни. Попасть в рай из бытия или низвергнуться в ад можно только за большой штраф g (смерть). Перемещаться по этим этажам (розовые линии) можно с низким штрафом h (h<<'g). Возвращение к жизни - реинкарнация - не наказывается штрафом. На втором этаже происходит только диагональное перемещение - мирские хлопоты, за которые мы пытаемся набрать наибольшее количество "очков" и получить наилучшее выравнивание (проход по красному контуру запрещён, он нужен для лучшего представления структуры объёмного графа). Проход по диагонали и получение бонусных баллов за удачное выравнивание осуществляется согласно начальной матрице весов. Ориентированный граф для построения локального выравнивания с линейными штрафами за гэпыРис. 12. Граф локального выравнивания с линейными штрафами. Собственная иллюстрация (участок графа 4х4) и готовая [2].
Для локального выравнивания нужно придать графу такие свойства, чтобы он позволил начать (закончить) выравнивание не с начала (в конце), а с любой вершины. Поэтому добавим стрелки с нулевой стоимостью от каждой вершины к концу и от начала до каждой вершины. То есть веса синих стрелок - 0, лиловых перпендикулярных переходов - линейный штраф за гэп g, вес прохода по диагонали (тёмно-фиолетовые рёбра графа) определяется матрицей весов. Матрица "весов дружелюбности" зрителей на концертеНачальные данные: трибуна актового зала МГУ на концерте классической музыки 21 апреля 2017 года. Расчёты и схема трибуны представлена на рисунке 13. Рис. 13. Рассадка людей на трибуне и расчёты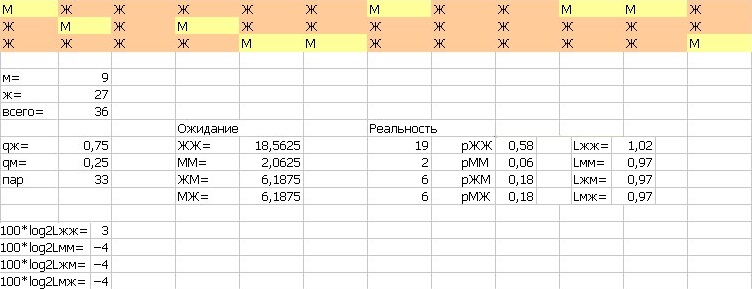
Табл. 5. Матрица весов дружелюбности
Скрипт для задания 2: локальное выравнивание фиксированного белка относительно других Источники [1] Алгоритм Смита — Ватермана, Википедия [2] Advanced Sequence Alignment |