Канальные белки - интегральные белки с полярными порами, осуществляющие пассивный транспорт через мембраны. В этом классе белков выделяют особую группу ионных каналов, осуществляющих облегченную диффузию ионов сквозь клеточные мембраны. Их классифицируют в соответствии с типом переносимых ионов. Наиболее важными считаются специфические ионные каналы, переносящие катионы Na+, K+, Ca2+ и анион Cl-. Значительно реже встречаются протонные каналы, а катионы магния переносятся посредством каналов, предназначенных для переноса других ионов. Другая классификация основывается на природе стимулов, влияющих на работу белков, на функциях и фармакологических свойствах. [7, 8]
Ионные каналы локализованы в цитоплазматической мембране клетки или в мембранах органелл; имеют симметричное кольцевое строение и состоят из 4, 5 или 6 структурно сходных доменов (субъединицы или отдельные белки). Характерно большое количество трансмембранных спиралей. В центре канала находится заполненная водой пора, через которую проходят ионы. В состоянии покоя ионы закрыты, открытие длится всего несколько миллисекунд и обычно регулируется различными стимулами: изменением потенциала мембраны, температуры, механическим напряжением или связыванием лиганда (в качестве лиганда могут выступать нейромедиаторы; наиболее распространенный тип). Перемещение ионов происходит в соответствии с градиентом концентрации и электрическим напряжением на мембране (электрохимический градиент). Стоит заметить, что в состоянии покоя обращенная к внешней среде часть цитоплазматической мембраны заряжена положительно, к внутренней - отрицательно. [7, 8, 9]
Рассматриваемый белок относится к катионным каналам транзиторного рецепторного потенциала (TRP) - крупной группе включающей 6 основных семейств ионных каналов плазматической мембраны, встречающихся у человека, животных и некоторых грибов.[1, 10]
Подсемейство TRPV названо так из-за ваниллоидных веществ, активирующих работу данных каналов. Как правило, связаны с молекулами капсаицина, восприимчивы к повышению температуры.
TRPV6 участвует в первой ступени абсорбции катионов кальция в пищеварительном тракте (проксимальная часть тонкой кишки), также был обнаружен в плаценте, почках, поджелудочной и предстательной железах. [10]
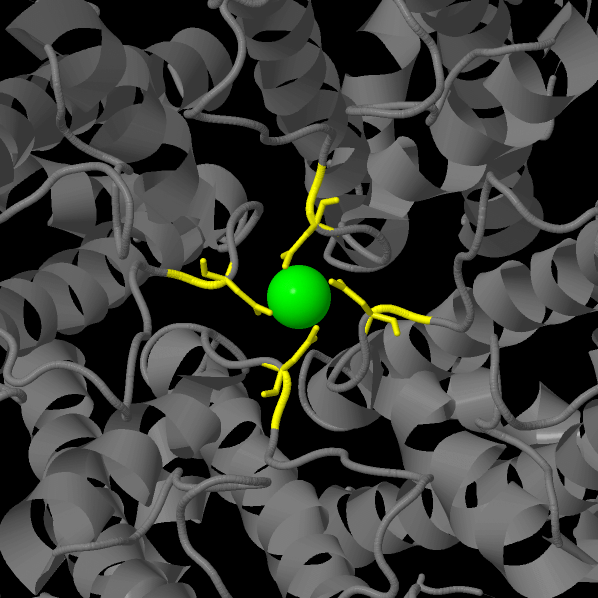
В этой работе рассматривается структура кристаллического TRPV6, взятого у Rattus norvegicus, состоящего из четырех субъединиц. Разрешение модели равно 3,247 Å. Как сообщают авторы, внутриклеточные домены ("юбка") вовлечены в аллостерическую регуляцию* процесса; специфичность трансмембранного домена связана с координацией катионов кальция кольцом боковых цепей аспартата в фильтре селективности (см. Рисунок 1). [1, 2, 4, 5, 10]
*Эффект ускорения процесса, возникающий при связывании молекулы-эффектора с белком вне активного центра последнего.
Рисунок 2, Рисунок 3. Взаимное положение лигандов и белковых субъединиц (зеленым обозначен ион кальция; cpk — DTB)
В состав комплекса входят всего два лиганда - катион кальция и 4 молекулы DTB (D-DESTHIOBIOTIN; C10H18N2O3). Взаимное положение лигандов иллюстируют рисунки 2 и 3.
Также в процессе работы мы обратились непосредственно к автору исследований, в результате которых была получена рассматриваемая структура TRPV6. Мы написали авторам статьи в общих словах о том, что знаем об этом белке, и спросили, не упустили ли мы какой-либо серьезной детали. С полученными статьями пока мы ознакомились лишь поверхностно, поскольку, судя по всему, ничего принципиально важного в нашем рассказе не упустили. На самом деле, интерес был скорее эмпирический, нежели нас действительно вынудила к этому работа. Это была наша первая попытка контакта с серьезной лабораторией и зарубежными авторами, и мы считаем, что она вполне успешна.
Представляем рекомендованные статьи для ознакомления:
Пептидная связь - вид ковалентной связи, возникающей в результате взаимодействия альфа-аминогруппы одной аминокислоты с альфа-карбоксильной группой другой. Данный вид связи в главным образом характеризует первичную структуру белка (полипептидная цепь). В таблице 1 представлены измерения некоторых рассмотренных нами пептидных связей, сделанные с помощью инструмента Measurment в программе для визуализации молекул Jmol.
Водородная связь - особый тип нековалентного взаимодействия, возникающий между атомом-донором, ассоциированным с водородом, и атомом-акцептором. Краткая характеристика водородных связей в рассмотренном белке, также составлянная с использованием программы Jmol, представлена в таблице 3.
Гидрофобные взаимодействия проявляются только в водных средах и обусловливаются способностью неполярных молекул образовывать между собой прочные ассоциаты в процессе мицеллообразования. Этим предопределяются возможность возникновения бислойных и многослойных биологических мембран, а также реализация конформационных переходов макромолекул белков.
Трехмерная структура поддерживается большим количеством внутримолекулярных нековалентных взаимодействий гидрофобной, природы, а также благодаря ионным взаимодействиям, дисульфидным мостикам, водородным связям и иногда солевым мостикам.
В трансмембранном домене рассматриваемого белка на поверхность выходят многочисленные гидрофобные альфа-спирали, обеспечивающие, по всей видимости, контакт с клеточной мембраной, а внутри канальной поры сосредоточены гидрофильные аминокислотные остатки.
Остальные домены также насыщены гидрофобными участками, но не так глобально, как это наблюдается в зоне соприкосновения с цитоплазматической мембраной.
Тип структур
Имена атомов
Длина связи(Å)
Угол N-H-O (°)
α-спираль
O(454SER)-N(458VAL)
3,6
160,6
310 спираль
O(467PHE)-N(470GLY)
3,2
154,2
β-лист
O(274SER)-N(267TRP)
3,6
156,0
Таблица 3. Характеристика водородных связей
Характерное расстояние между соседними не связанными ковалентно атомами в белке - 5,336.
Между некоторыми соседними атомами может поместиться молекула воды. Такое возможно, например, между [LEU]399:A.CD1 и [PHE]433:A.CD1; между остальными представленными в таблице 4 парами атомов - нет.
Это обусловлено тем, что ван-дер-ваальсовый радиус атома кислорода (равный 1,4 Å), взятый за ван-дер-ваальсовый радиус молекулы воды, превосходит половину значения расстояния между соседними не связанными ковалентно атомами во всех описанных случаях, кроме последнего, где полурасстояние между атомами составляет 1,45 Å.
Расстояние между соседними атомами рассчитано с учетом их ван-дер-ваальсовых радиусов (у углерода он равен 1,85 А).
Атом 1
Атом 2
Расстояние
Полурасстояние (с учетом ван-дер-ваальсовых радиусов)
Солевые мостики (солевые связи; ионные связи) - нековалентные связи, возникающие в белке между разноименно заряженными радикалами аминокислотных остатков, располагающихся в гидрофобном ядре. Например, между группами -COO- (аспагариновая или глутаминовая кислоты) и -NH3+ (лизин). Солевые связи могут быть внутри- и межцепочечными [11]. Солевые мостики стабилизируют структуру белка, хотя внутренняя ионная связь встречается редко, потому что большинство заряженных аминокислот лежат на поверхности белка, такие связи являются мощными электростатическими притяжениями, которые могут быть близки по прочности к ковалентным связям[12, 13].
Для поиска солевых мостиков были выделены полярные остатки аминокислот и раскрашены в соответствии с зарядом:
restrict polar and not backbone
select acidic
color red
select basic
color cyan
Визуально подходившие пары были отобраны. Для проверки мы измерили расстояния между заряженными группировками (3,24 Å, 4,94 Å, 2,59 Å) и обратили внимание на положение частиц в молекуле (все они оказались в гидрофильной части вблизи границы молекулы вне трансмембранного домена).
Стэкинг-взаимодействия (π -стэкинг) - тип нековалентных взаимодействий, возникающий между ароматическими кольцами. Такие взаимодействия играют большую роль в укладке нуклеиновых оснований в молекулах ДНК, РНК и в формировании структуры белков.
Чтобы найти стекинг-взаимодействия, была выполнена команда restrict aromatic and not backbone. Выделив боковые цепи ароматических аминокислот контрастным цветом, мы легко нашли несколько примеров стекинг-взаимодействий. Нетрудно увидеть, что они удовлетворяют критериям на минимальные расстояния между кольцами, изложенным в [14]. (3,83Å, 3,44 Å)
ЛИГАНД-БИОМОЛЕКУЛЯРНЫЕ КОНТАКТЫ
Дестиобиотин является синтетическим метаболитом, который имитирует действие биотина (водорастворимый витамин группы В) на экспрессию генов и, таким образом, обладает биотиноподобной активностью. У млекопитающих биотин служит коферментом для карбоксилаз, таких как пропионил-КоА-карбоксилаза [15]. О роли DTB в данном белке пока не известно.
Дестибиотин связан с нашим белком как с помощью гидрофобных контактов, так и водородными связями (заметна только одна, вероятно, есть и вторая. Заряд при физиологическом pH DTB равен –1, pKa - 4,21. Возможно, белок закристаллизовался таким образом, что в момент внутреннего колебания структуры одна из двух равноценных по идее связей DTB азотами из белка оказалась слишком длинной чтобы JMol считал ее связью) [1]. Одна молекула присутствует в каждой белковой субъединице. (см. апплет "Show DTB-ligand bonding"; см. Рисунок 4)
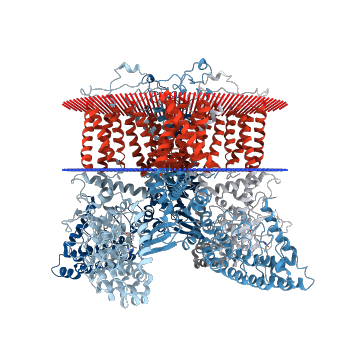
Рисунок 4. Изображение белка, погруженного в мембрану (www.rcsb.org)
Рисунок 5. Изображение молекулы дестибиотина
Рисунок 6. Структурная формула дестибиотина
ВЫВОДЫ
В результате изучения структуры молекулы, доступной в банке PDB, и других источников удалось составить атлас контактов. В молекуле нет дисульфидных мостиков, а основные типы связей в третичной структуре — водородные связи, ионные пары (солевые мостики), стекинг- и гидрофобные взаимодействия. Примечательно, что в местах контакта с мембраной клетки поверхность молекулы более гидрофобна, чем на частях, контактирующих с цитозолем и с межклеточным пространством, и даже чем внутри самого канала.
В белке есть два типа низкомолекулярных лигандов: DTB и ион кальция. DTB удерживается в структуре в основном гидрофобно, а кальций, проходя сквозь канал, может координировать аспартатное кольцо. В структуре из банка PDB ион металла показан хелатированным [3, 6].
** Обращаем ваше внимание, что не все скрипты корректно работают в апплет-версии.
Личный вклад
Мария Кузнецова исследовала контакты белок-лиганд (DTB), пептидные и водородные контакты в белке; написала скрипты к демонстрации вышеперечисленных данных и сделала необходимые измерения некоторых параметров пептидных и водородных связей; частично нашла информацию про лиганд и некоторые контакты.
Руслан Салимгареев участвовал в подборе литературы, нашёл примеры ионных взаимодействий и стекинга, написал скрипт, демонстрирующий стекинг; участвовал в финальном редактировании отчета.
Ирина Боровикова оформила html-страницу, собрала материалы и написала обзорную часть; участвовала в написании других текстовых элементов; участвовала в координации работы команды; участвовала в обсуждении и создании скриптов, редактировала скрипты, в т.ч. для работы в апплет-версии; участвовала в финальном редактировании отчета.
Елизавета Силинг перевела страницу на английский язык, участвовала в поиске статей для обзора, написала письмо заведующему лабораторией Sobolevsky Lab, исследовала гидрофобные взаимодействия и писала скрипт к их демонстрации, исследовала плотность расположения атомов в гидрофобном ядре; написала скрипт, описывающий ионные взаимодействия.
Ссылки на источники
[1] http://www.rcsb.org/structure/5IWK
[2] https://cyberleninka.ru/article/v/sovremennye-predstavleniya-ob-ionnyh-kanalah-i-kanalopatiyah-obzor-literatury
Салмина А.Б., Шнайдер Н.А., Михуткина С.В, “Современные представления об ионных каналах и каналопатиях (обзор литературы)”
[3] http://www.iephb.ru/wp-content/uploads/dissertation44.pdf
Коркош В.С., “Молекулярное моделирование потенциал-управляемых натриевых каналов эукариот и их взаимодействия с лигандами”, 2015
[4] https://sobolevskylab.org/structure-and-gating-mechanism-of-the-transient-receptor-potential-channel-trpv3/
Appu K. Singh, Luke L. McGoldrick, Alexander I. Sobolevsky, “Structure and gating mechanism of the transient receptor potential channel TRPV3”, Nature Structural & Molecular Biology, 08.20.2018
[5] https://doi.org/10.1038/nature17975
Saotome, K., Singh, A. K., Yelshanskaya, M. V., & Sobolevsky, A. I. (2016). Crystal structure of the epithelial calcium channel TRPV6. Nature, 534(7608), 506–511.