Channel proteins are integral proteins with polar pores that carry through passive transport through membranes. A special group of ion channelsin this class of proteins facilitate the light diffusion of ions through cell membranes. They are classified according to the type of ions they transfer. Specific ion channels carrying the Na+, K+, Ca2+ cations and the Cl- anion seem the most important. Vastly less frequently, proton channels are found, and magnesium cations are transferred by means of channels intended to transfer other ions. Another classification is based on the nature of the stimuli that affect the protein work, on functions and pharmacological properties. [7, 8]
Ion channels are localized in the cytoplasmic membrane of the cell or in the membranes of organelles; have a symmetrical ring structure and consist of 4, 5, or 6 structurally similar domains (subunits or individual proteins). A large number of transmembrane helices is typical. In the center of the channel there is a pore filled with water through which ions pass. At resting state, ions are closed, the opening lasts only a few milliseconds and is usually regulated by various stimuli: changes in membrane potential, temperature, mechanical stress, or ligand binding (the neurotransmitters may be the ligand; the most common type). The migration of ions occurs in accordance with the concentration gradient and the voltage on the membrane (electrochemical gradient). It is worth noting that in the state of rest, part of the cytoplasmic membrane that is turned to the external medium is positively charged, to the internal part - negatively. [7, 8, 9]
The protein under consideration belongs to the cationic channels of the transient receptor potential (TRP) - a large group that comprises 6 main families of plasma membrane ion channels found in humans, animals, and some fungi. [1, 10]
The subfamily TRPV is named so because of the vanilloid substances that activate the work of these channels. Generally TRPVs are associated with capsaicin molecules that are susceptible to the temperature rise.
TRPV6 is involved in the first step absorption of calcium cations in the digestive tract (proximal part of the small intestine) and has also been found in the placenta, kidneys, pancreas and prostate glands. [10]
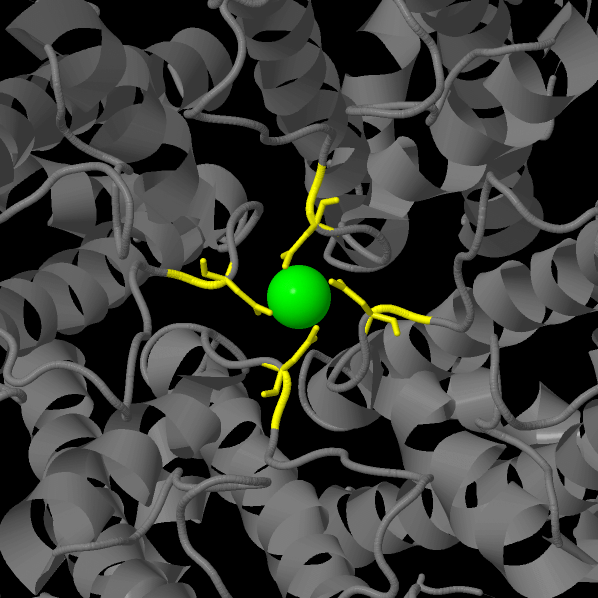
This work is considered to discuss the structure of crystalline TRPV6 consisting of four subunits taken from Rattus norvegicus. The resolution of the model is 3.247 Å. According to the authors of the article, intracellular domains ("skirt") are involved in the allosteric regulation* of the process. The specificity of the transmembrane domain is associated with the coordination of calcium cations by the ring of aspartate side chains in the selectivity filter (see Figure 1). [1, 2, 4, 5, 10]
*The effect of the process acceleration that occurs when the effector molecule binds to a protein outside its active center.
Figure 1. Aspartate ring (calcium ion is green, aspartic acid residues are yellow)
The complex consists of only two ligands - a calcium cation and 4 molecules of DTB (D-DESTHIOBIOTIN; C10H18N2O3). The mutual position of the ligands is illustrated by figures 2 and 3.
Figure 2, Figure 3. The mutual position of the ligands and protein subunits (calcium ion is green; cpk — DTB)
During the process of work, we have also written directly to the author of the research, as a result of which the structure of TRPV6 was obtained. In Figure 4, you can see the text of the response letter.
Figure 4. Response from A. I. Sobolevsky
We represent the articles for review that were recommended:
Peptide bond is a type of covalent bond resulting from the interaction of the alpha-amino group of one amino acid with the alpha-carboxyl group of another one. This type of bond mainly characterizes the primary structure of the protein (polypeptide chain). Table 1 represents the measurements of some of the peptide bonds that we considered, using the Measurment tool in the program for molecules visualization of Jmol.
A hydrogen bond is a special type of non-covalent interaction that occurs between a donor atom associated with a hydrogen and an acceptor atom. A brief characteristic of the hydrogen bonds in the considered protein, that was also compiled using the Jmol program, is represented in Table 3.
Hydrophobic interactions occur only in aqueous media and are determined by the ability of non-polar molecules to form strong associates between themselves in the process of micelle formation. This predetermines the possibility of the emergence of bilayer and multilayer biological membranes, as well as the realization of conformational transitions of protein macromolecules.
The three-dimensional structure is supported by a large number of intramolecular non-covalent interactions of hydrophobic nature, as well as by ionic interactions, disulfide bridges, hydrogen bonds and sometimes salt bridges. Hydrophobic interactions are the most important because they are supposed to be responsible for the large amount of free binding energy that is observed during enzyme-substrate interactions.
In the transmembrane domain of the protein that has been considered, numerous hydrophobic alpha helixes appear on the surface, apparently providing contact with the cell membrane, and hydrophilic amino acid residues are concentrated inside the channel pore.
The remaining domains are also saturated with hydrophobic sites, but not as fully as it is observed in the zone of contact with the cytoplasmic membrane.
Type of structure
Atom names
Bond length (Å)
Angle N-H-O (°)
α-helix
O(454SER)-N(458VAL)
3,6
160,6
310 helix
O(467PHE)-N(470GLY)
3,2
154,2
β-sheet
O(274SER)-N(267TRP)
3,6
156,0
Table 3. Characteristics of hydrogen bonds
The characteristic distance between adjacent unrelated covalently unrelated atoms in a protein is 5.336.
Between some of the adjacent atoms an H2O molecule can fit. This is possible, for example, between [LEU]399:A.CD1 and [PHE]433:A.CD1, but not between the remaining pairs of atoms represented in the table.
This happens because Van der Waals radius of the oxygen atom (equal to 1.4 angstroms), taken as the Van der Waals radius of the water molecule, exceeds half the distance between adjacent atoms that are not covalently connected in all the described cases, except the last, where the half-distance between atoms is 1.45 angstroms.
The distance between neighboring atoms is calculated their van der Waals radii (for carbon it is equal to 1.85 A) taken into account.
Atom 1
Atom 2
Distance
Half-distance (after calculating Van der Waals radii)
Does H2O molecule fit?
[PHE]152:A.CD1
[PHE]169:A.CZ
3.8728158
0.1228158
Нет
[PHE]152:A.CE2
[GLN]118:A.CB
4.145851
0.445851
Нет
[PHE]152:A.CB
[GLY]163:A.O
4.0248337
0.3248337
Нет
[PHE]152:A.CZ
[GLN]118:A.CG
4.007123
0.307123
Нет
Table 4. Characteristic of hydrophobic interactions
Ionic bonds are non-covalent bonds that occur in a protein between oppositely charged radicals of amino acid backbones located in a hydrophobic core. For example, between the -COO- (aspagaric or glutamic acid) and -NH3 + (lysine) groups. Ionic bonds can be intra- and interchain [11]. Ionic bonds stabilize the structure of the protein. Although the internal ionic bond is rare, because most of the charged amino acids are located on the surface of the protein, such bonds are a powerful electrostatic attraction, which can be similar to covalent bonds in strength [12, 13].
To search for the ionic bonds, polar amino acid residues were isolated and colored according to their charge:
restrict polar and not backbone
select acidic
color red
select basic
color cyan
Pairs that have matched visually were selected. To check, we measured the distances between charged groups (3.24 Å, 4.94 Å, 2.59 Å) and paid attention to the position of the particles in the molecule (all of them were in the hydrophilic part near the molecular boundary outside the transmembrane domain).
Stacking interactions (π-stacking) are a type of non-covalent interactions that occur between aromatic rings. Such interactions play an important role in the folding of nucleic bases in DNA molecules, RNA, and in the formation of protein structures.
To find stacking interactions, the “restrict aromatic and not backbone” command was executed. We have easily found several examples of stacking interactions by highlighting the aromatic amino acid side chains in a contrasting color. It is easy to notice that they satisfy the criteria for the minimum distances between the rings described in [14]. (3.83Å, 3.44 Å)
LIGAND-BIOMOLECULAR CONTACTS
Destobiotin is a synthetic metabolite that imitates the action of biotin (a water-soluble vitamin of group B) on gene expression and, thus, has biotin-like activity. In mammals, biotin serves as a coenzyme for carboxylase, such as propionyl-CoA-carboxylase [15]. The role of DTB in this protein is not known yet.
Destibiotin is associated with our protein using both hydrophobic contacts and hydrogen bonds (only one is noticeable, probably the second. The physiological charge (at physiological pH) of DTB is -1, pKa equals 4.21. It is possible that the protein crystallized in such a way that The structure of one of the two equivalent DTB bonds with nitrogen from the protein turned out to be too long for JMol to regard it as a bond) [1]. One molecule is present in each protein subunit. (see the applet "Show DTB-ligand bonding"; see Figure 5)
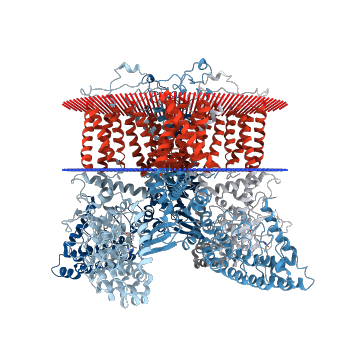
Figure 5. Image of protein immersed in the membrane (www.rcsb.org)
Figure 6. Image of the Destibiotin Molecule
Figure 7. Image of the destibiotin structural formula
CONCLUSIONS
As a result of studying the structure of the molecule available in the PDB bank and other sources, it has become possible to compile an atlas of contacts. There are no disulfide bridges in the molecule, and the main types of bonds in the tertiary structure are & nbsp; & mdash; hydrogen bonds, ion bonds, stacking and hydrophobic interactions. It is noteworthy that in places of contact with the cell membrane, the surface of the molecule is more hydrophobic than in the parts in contact with the cytosol and the extracellular space, and even within the channel itself.
There are two types of low molecular weight ligands in the protein: DTB and calcium ion. DTB is retained in the structure mainly by hydrophobic interactions, and calcium, while passing through the channel, can coordinate the aspartate ring. In the structure from the PDB bank, the metal ion is shown to be chelated [3, 6].
** Please note that not all scripts work correctly in the applet version.
Personal contribution
Maria Kuznetsova has studied the contacts of the protein-ligand (DTB), peptide and hydrogen contacts in a protein; written scripts to demonstrate the data above and made the necessary measurements of some parameters of peptide and hydrogen bonds; partially found information about the ligand and some of the contacts.
Ruslan Salimgareev has participated in the selection of literature, found examples of ionic interactions and stacking, and written the script that demonstrates stacking.
Irina Borovikova has designed the html-page, collected materials and written the review part, investigated the work of the channel systems; participated in writing of other text elements; participated in coordination of the work of the team; participated in discussion and creation of scripts, edited scripts and also made them work in the applet version, participated in the final editing of the report.
Elizaveta Siling has translated the page into English, participated in the search for articles for review, written a letter to the head of the laboratory of Sobolevsky Lab, investigated hydrophobic interactions and written a script for their demonstration, investigated the density of atoms in the hydrophobic core; written a script describing ionic interactions.
References
[1] http://www.rcsb.org/structure/5IWK
[2] https://cyberleninka.ru/article/v/sovremennye-predstavleniya-ob-ionnyh-kanalah-i-kanalopatiyah-obzor-literatury
Салмина А.Б., Шнайдер Н.А., Михуткина С.В, “Современные представления об ионных каналах и каналопатиях (обзор литературы)”
[3] http://www.iephb.ru/wp-content/uploads/dissertation44.pdf
Коркош В.С., “Молекулярное моделирование потенциал-управляемых натриевых каналов эукариот и их взаимодействия с лигандами”, 2015
[4] https://sobolevskylab.org/structure-and-gating-mechanism-of-the-transient-receptor-potential-channel-trpv3/
Appu K. Singh, Luke L. McGoldrick, Alexander I. Sobolevsky, “Structure and gating mechanism of the transient receptor potential channel TRPV3”, Nature Structural & Molecular Biology, 08.20.2018
[5] https://doi.org/10.1038/nature17975
Saotome, K., Singh, A. K., Yelshanskaya, M. V., & Sobolevsky, A. I. (2016). Crystal structure of the epithelial calcium channel TRPV6. Nature, 534(7608), 506–511.