В ходе данной работы используется база данных метаболических путей проекта KEGG, доступная по адресу: http://www.genome.jp/kegg/pathway.html. Анализу подверглись метаболические пути неканонической аминокислоты глутатиона.
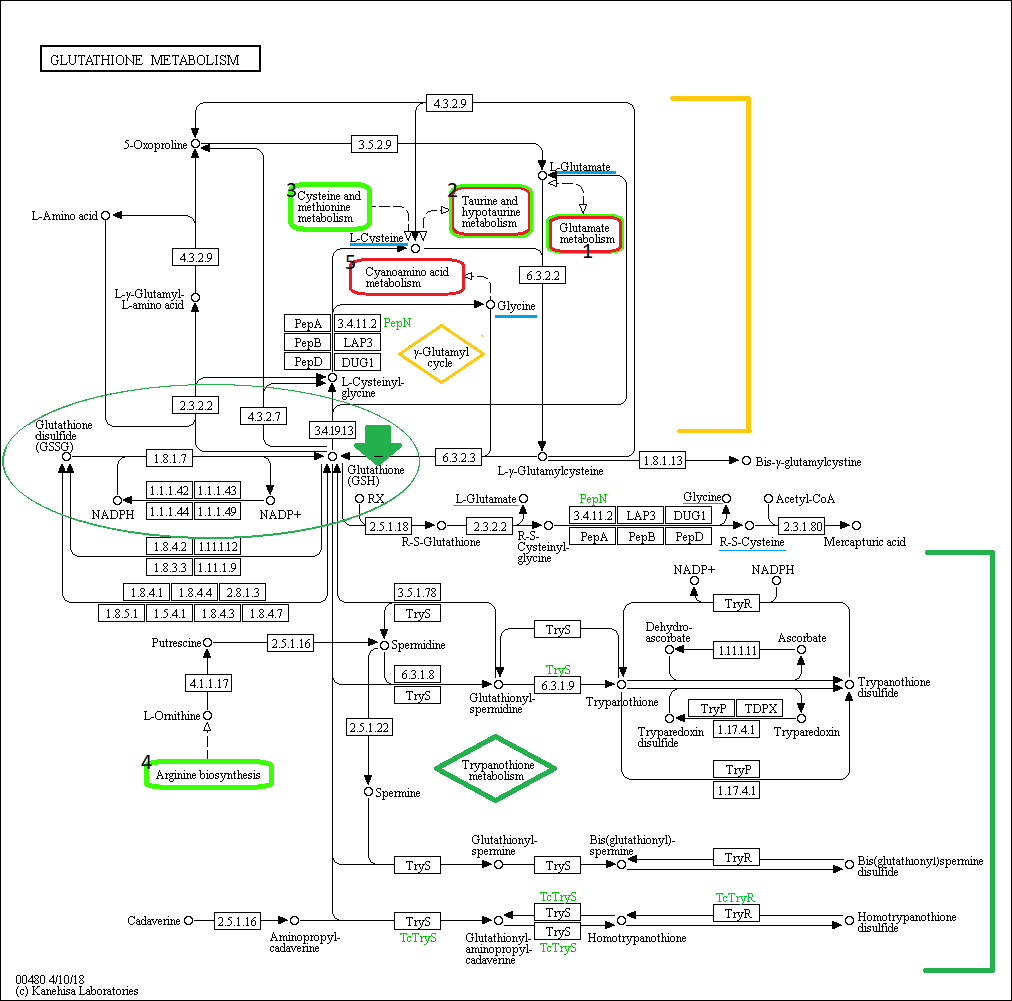
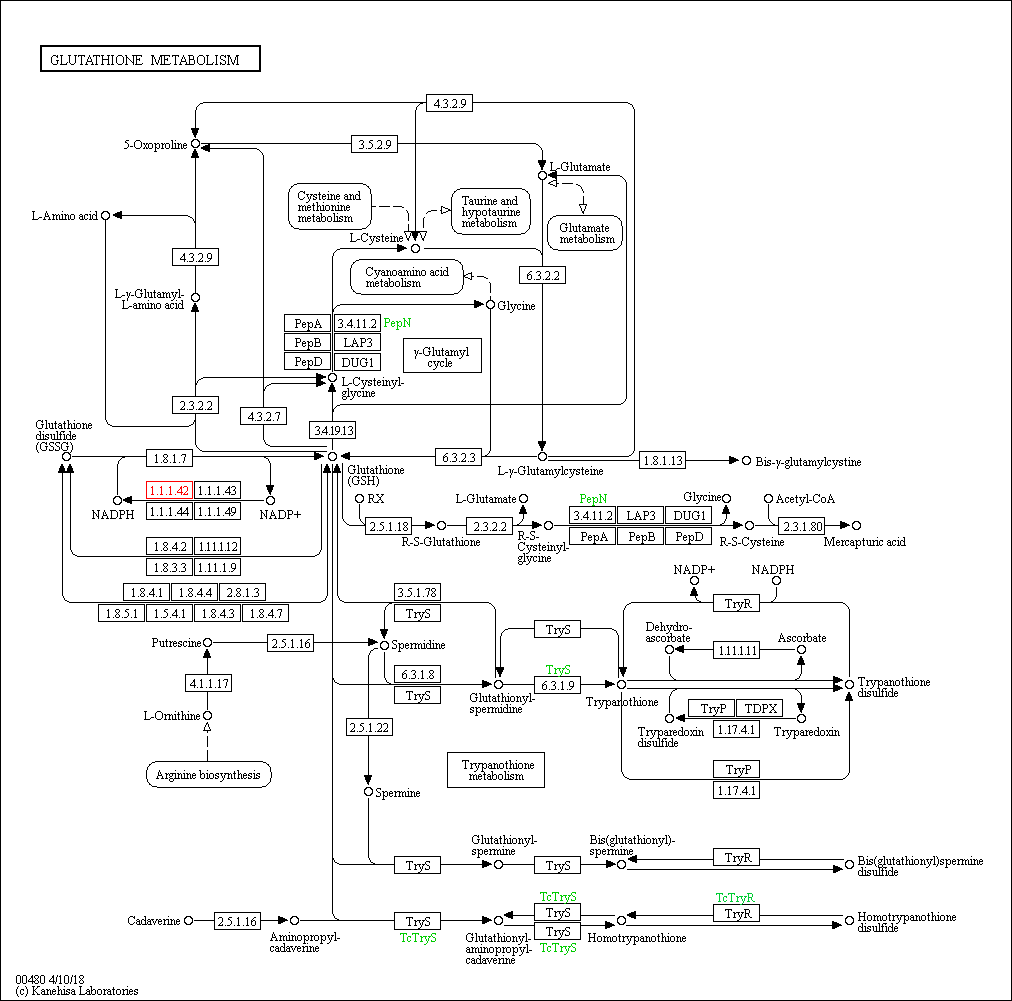
Ниже приведена карта исследуемого пути метаболизма глутатиона:
| Рисунок 1. Метаболизм глутатиона, KEGG id: map00480, доступен в б.д. KEGG по ссылке http://www.genome.jp/kegg-bin/show_pathway?map00480 Зелёным овалом показана основная редокс-буферная система млекопитающих, Глутатион - GSH(обозначен стрелочкой) и его дисульфидная окисленная форма - GSSG, осуществляющая защиту от окислительного стресса в клетках. Синим подчеркнуты исходные для синтеза аминокислоты. Жёлтой скобкой - гамма-глутамиловый цикл, зелёной скобкой - метаболизм трипанотиона. Сопряжённые метаболические пути обозначены цифрами и цветными овалами: овалы красного цвета обозначают метаболическе пути, заимствующие вещества из метаболического пути GSH, овалы салатового цвета обозначают метаболическе пути, донирующие некоторые метаболиты. |
Глутатион (2-амино-5-{[2-[(карбоксиметил)амино]-1-(меркаптометил)-2-оксоэтил]амино}-5-оксопентаноевая кислота, трипептид ?-глутамилцистеинилглицин, далее в тексте - GSH) является наиболее важным низкомолекулярным антиоксидантом, синтезируемым в клетках. Сульфгидрильная группа (-SH) в его составе обеспечивают удаление пероксидов и многих ксенобиотических соединений; однако GSH также участвует в регуляции клеточного цикла, в реакциях детоксификации, маркируя токсические вещества в процессе их биотрансформации с целью выведения из организма, и в синтезе лейкотриенов. Взаимодействие GSH с глутатион-S-трансферазой инициирует реакции конъюгации и восстановления, катализируемые в цитозоле, микросомах и в митохондриях [1].
Метаболизм GSH можно кластеризовать вокруг двух несмежных групп реакций: гамма-глутамиловый цикл и метаболизм трипанотионина - который далее будем обозначать Try. Try является димеризованной формой GSH, содержащей две молекулы GSH, соединённых компоновщик - spermidine (спермидином), который поставляется из сопряжённого с метаболизмом GSH биосинтезом аргинина в форме L-орнитина. Try наравне с GSH участвут в поддержании редокс-баланса в клетке [2].
В организме человека GSH синтезируется из аминокислот L-цистеина, L-глутамата и глицина, обеспечивая сульфгидрильная группой в составе последнего обеспечивает наиболее важные функциии GSH по поддержанию окислительно-восстановительного баланса. Связь между метаболическими путями исходных веществ осуществляется в метаболизме GSH непосредственно через эти вещества - см. схему.
Синтез в организме происходит в гамма-глутамиловом цикле в две ATP-зависимые стадии: 1)синтез ?-глутамилцистеина из глутаминовой кислоты и цистеина ферментом ?-глутамилцистеинсинтетазой; 2)присоединение остатка глицина к С-концевой группе ?-глутамилцистеина. Первая стадия является лимитирующей в процессе синтеза [3].
Ниже указаны некоторые конченые и начальные вещества в метаболизме: в данном контексте это, соответственно, вещества, стоящие в начале линейной цепочки реакций, которые в рамках этого метаболизма нельзя получить из каких-либо других, кроме как обратимой реакцией, и вещества, стоящие в конце той же самой линейной цепочки реакций, из которых в рамках данного метаболизма нельзя получить ни одно другое, кроме как обратимой реакцией. В случае обратимых реакций записан только один вариант той. В таблице не включён гамма-глутамил цикл.
| Таблица 1.Начальные и конечные вещества пути | |
|---|---|
| Начальные | Конечные |
| Кадаверин | Гомотрипанотиона дисульфид |
| L-орнитин | Би(глутатионил)сперминдисульфид |
| L-орнитин | Трипанотионина дисульфид |
| GSH | Меркаптуриновая кислота |
| GSH | GSSG |
| GSH | Гомотрипанотиона дисульфид |
| GSH | Би(глутатионил)сперминдисульфид |
Из доступных в базе KEGG организмов были выбраны метаболические пути GSH для Escherichia coli/Кишечной палочки, Methanocaldococcus jannaschii, Pan troglodytes/Обыкноыенного шимпанзе. По базе данных Taxonomy библиотеки NCBI ниже представлена таксономия для данных организмов.
| Таблица 2.Таксономия выбранных организмов | |||||
|---|---|---|---|---|---|
| Domain | Phylum | Class | Genus | Species | |
| Eukaryota | Chordata | Mammalia | Pan | Pan troglodytes | |
| Archaea | Euryarchaeota | Methanococci | Methanocaldococcus | Methanocaldococcus jannaschii | |
| Bacteria | Proteobacteria | Gammaproteobacteria | Escherichia | Escherichia coli (str. K-12 substr. MG1655) | |
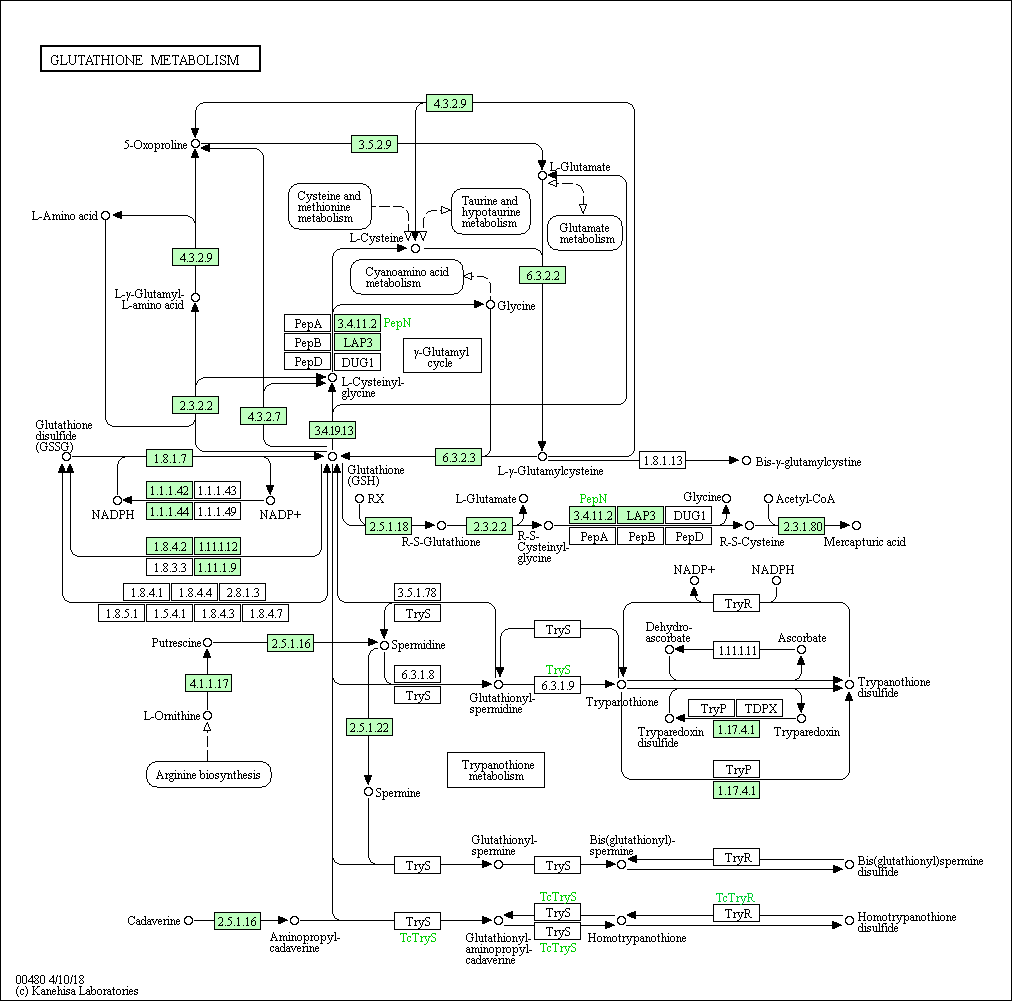
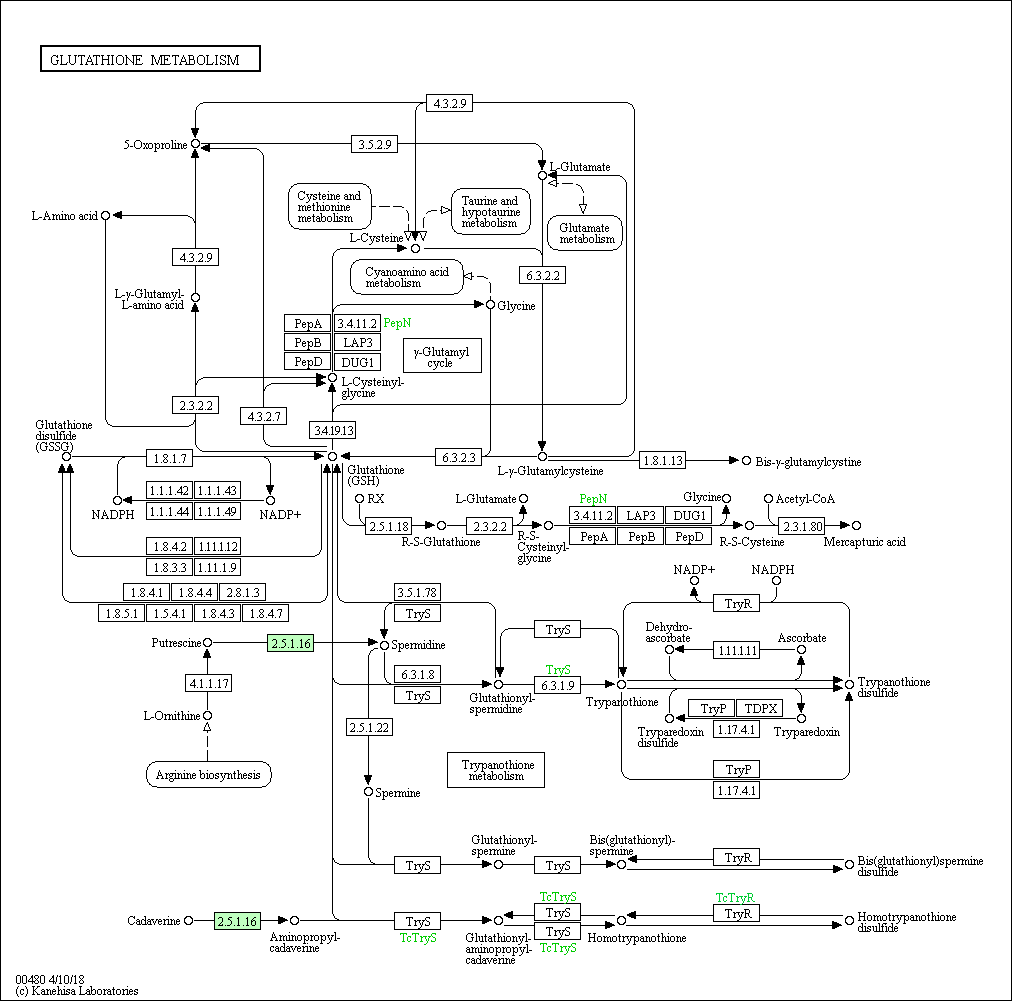
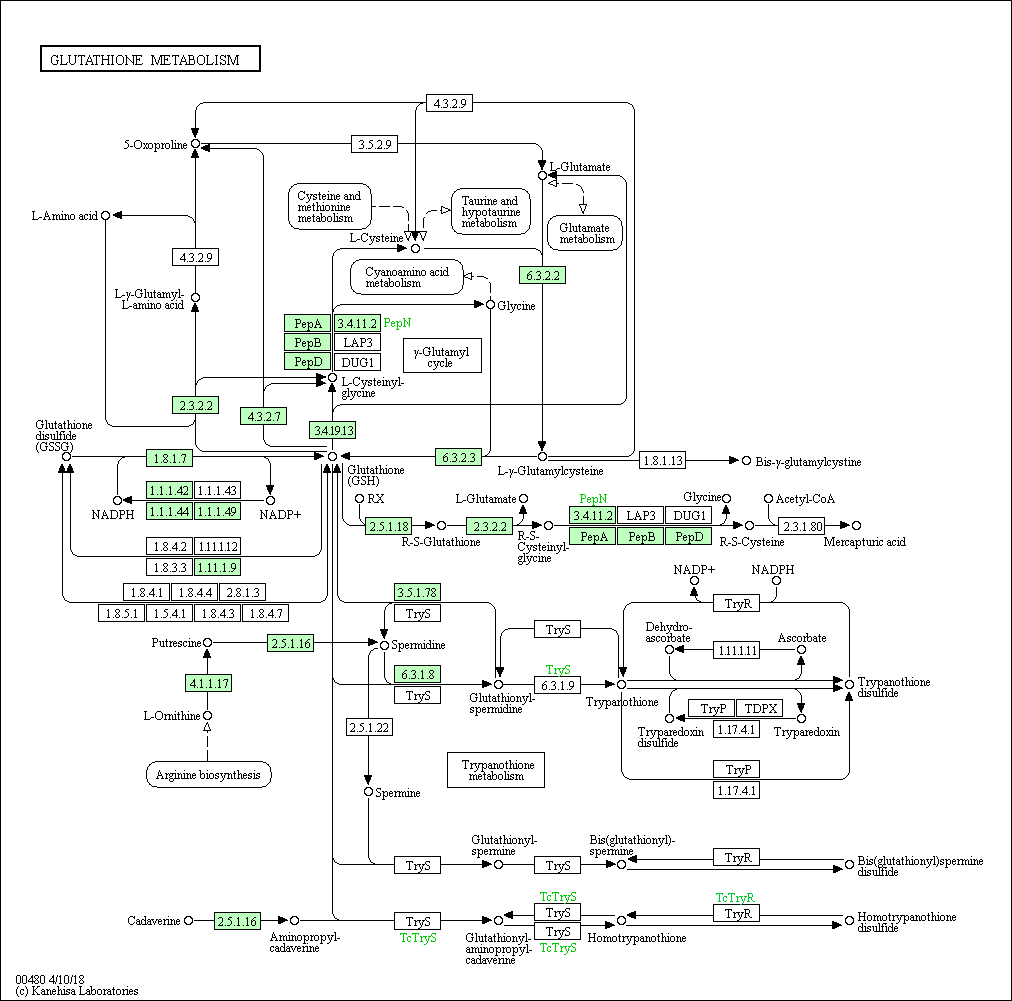
На основании данных метаболических карт мы можем судить о возможности протекания реакций метаболических путей GSH в этих организмах.
Полнее всего метаболизм GSH представлен у шимпанзе, где полностью протекают гамма-глутамиловый цикл и метаболизм трипанотиона, реализуются GSH-GSSG переход, синтез Bis-γ-glutamylcystine, Mercapturic acid. Линейная цепочка из исходного пути метаболизма кадаверин --> гомотрипанотионина дисульфид сохраняется. Трипанотионовый метаболизм распадается, остаётся возможность синтеща дисульфида трипанотиона из трипанотиона, последний производит циклическое превращение Tryparedoxin disulfide - Tryparedoxin через фермент ribonucleoside-diphosphate reductase (EC:1.17.4.1) и, отдельно, синтещ спермина из L-орнитина. В этом же организме представлено большиснтво ферментов.
У E.coli γ-глутамиловый цикл остался не полностью, он представляет из себя несколько разветвлённых реакций, продуктами GSH в котором служат 5-оксопролин и L-γ-гулатмил-L-аминокислота, непереходящи друг в друга. Циклическими остаются переходы GSH->L-цистеинил-глицин->L-цистеин->L-глутамилцистеин->GSH и GSH->L-цистеинил-глицин->глицин->GSH. Редокс-буфер GSH-GSSG так же сохраняет свою функцию, потому как этот переход в организме осуществим. Реализуются так же синтезы Bis-γ-glutamylcystine, Mercapturic acid, но в рамках метаболизма трипанотиона возможна теперь более полная линейная цепочка: от L-орнитина из аргининового биосинтеза до трипанотионина (синтез обрывается ранее, нежели чем в предыдущем случае), с возможностью (чего так же не было в прошлом случае) перехода промежуточного глутитаонил-спермицина в GSH. Линейная цепочка из исходного пути метаболизма кадаверин --> гомотрипанотионина дисульфид так же сохраняется.
У археи Methanocaldococcus jannaschii метаболизм GSH можно считать не представленным. У этого организма, как и у всех предыдущих, представлена линейная цепочка кадаверин --> гомотрипанотионина дисульфид характерная для изначального метаболизма, когда как сохранившийся обрывок синтеза трипанотионина: putrescine --> спермидин не имеет в своём составе краевых точек исходного пути. То же самое для γ-глутамилового цикла: фермент PepN, представленный в данном организме, способен опосредовать единственную реакцию: распад цистеинил-глицина на глицин и цистеин.
Выводы:
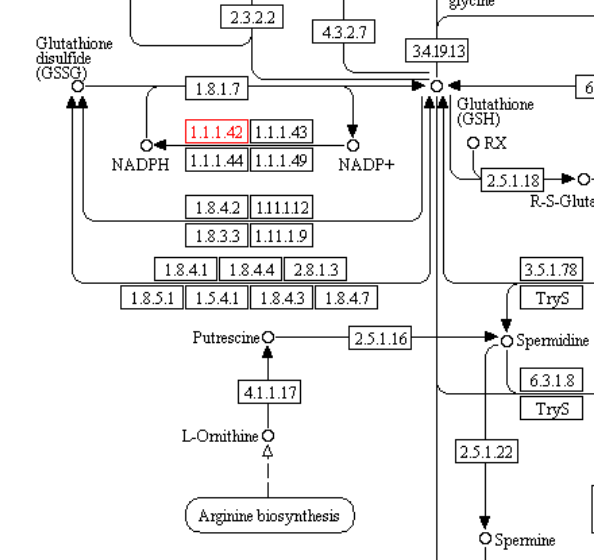
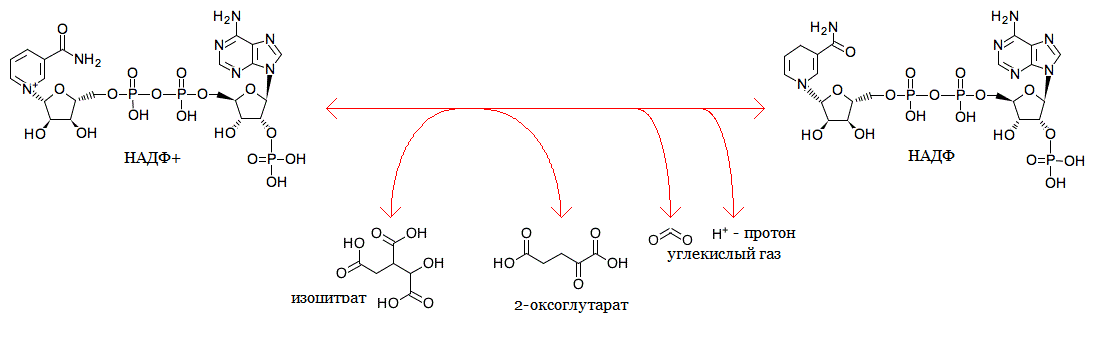
Данная реакция представлена в базе данных KEGG под идентификатором R00267 или по ссылке: http://www.genome.jp/dbget-bin/www_bget?rn:R00267 В метаболизме GSH реакция происходит на стадии перехода восстановленного GSH в его окисленную форму - GSSG и осущствляется ферментом изоцитрат-дегидрогеназой (EC 1.1.1.42).
 |  |
На главную страницуВернуться назад
©Solonovich Vera,2017