Рекоснтрукция филогенетического дерева представителей Metazoa по последовательностям митохондриальной рРНК
Реконструкция дерева по последовательностям РНК 12S-субъединицы митохондриальной рибосомы
Для реконструкции филогенетического дерева были взяты те же организмы что и ранее, за несколькими исключениями, для которых отсуствовали полные геномы митохондрий: вместо Sarcophyton glaucum был взят Sarcophyton minusculum (9CNID), вместо Caenorhabditis briggsae - Caenorhabditis nigoni (9PELO). Также поскольку длина последовательности митохондриальной рРНК у Paraspadella gotoi оказалась заметно меньше, чем у остальных организмов (менее 550 н. по сравнению с 700-1200 у остальных), дополнительно была взята рРНК родственного вида Spadella cephaloptera (9BILA).
Полученные нуклеотидные последовательности были выровнены при помощи алгоритма MAFFT и при помощи программы fastme было построено дерево (Рис. 1). Для определения эволюционных расстояний использовалась метрика Тамуры-Неи (TN93), также игнорировались все столбцы выравнивания, содержащие гэпы, поскольку в полученном выравнивании более половины столбцов содержало гэпы почти во всех строках.
fastme -i 12S_aligned.phy -dT -r -o 12S_T.phy
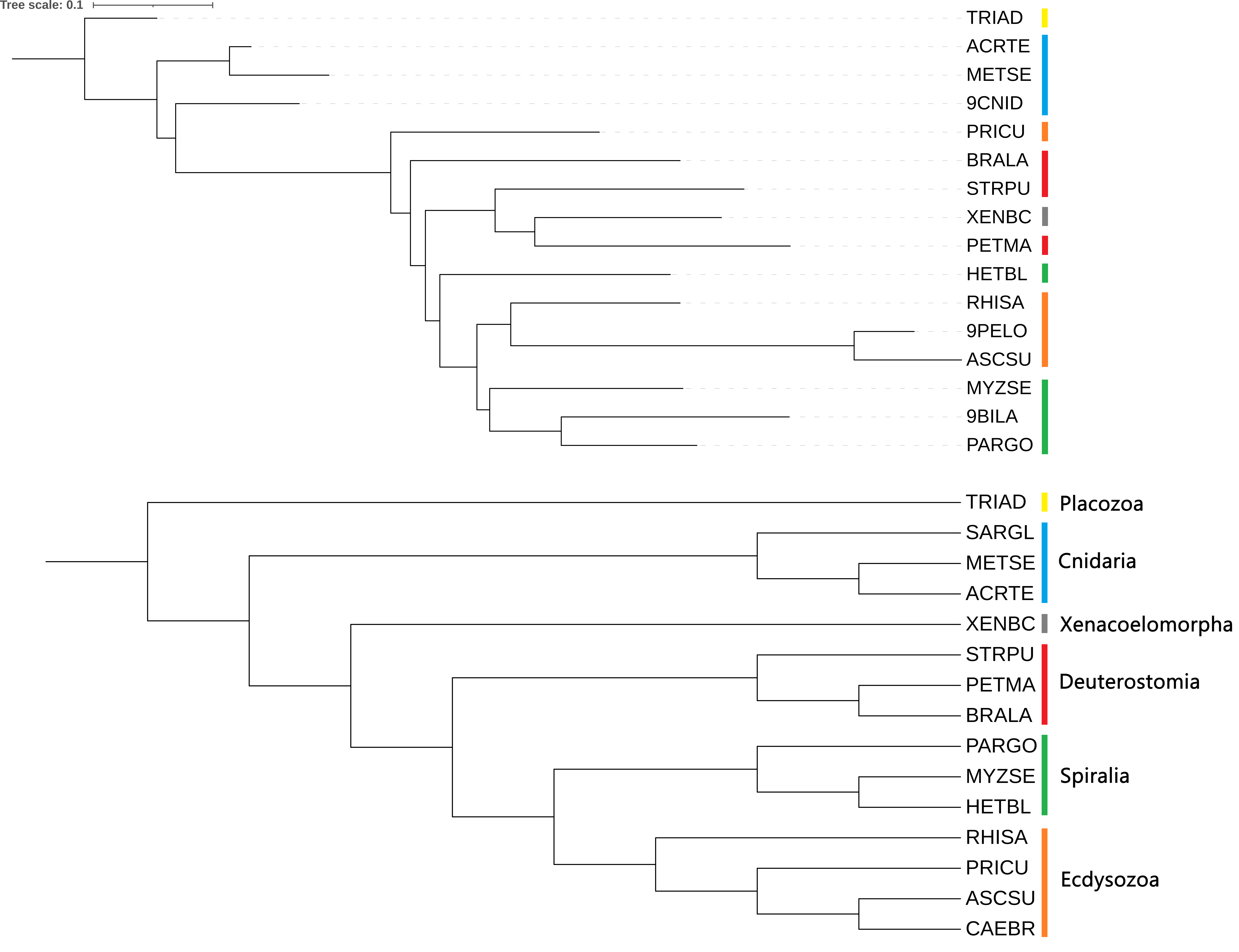
Как можно видеть, реконструкция филогении по РНК 12S-субъединицы митохондриальной рибосомы дает более расходящиеся с ожидаемыми результаты, нежели реконструкция по аминокислотным последовательностям цитохрома B. В данном случае ни одна крупная монофилетическая группа не сохранилась, но можно видеть что по прежнему две нематоды (ASCSU и 9PELO) являются ближайшими родственниками, а также таковыми оказались виды из группы Spadellidae (PARGO и 9BILA).
Укоренение дерева во внешнюю группу
Поскольку рассматриваемы набор представителей Metazoa включает представителей сильно отличных друг от друга групп, выбрать группу из Metazoa, которая являлась бы внешней для всех выбранных представителей можно было бы только принимая на веру одну из гипотез об эволюционных отношениях между самыми крупными группами Metazoa (такими как Porifera, Ctenophora, Placozoa и др.). Поэтому в качестве внешней группы был выбран представитель Choanoflagellata Monosiga brevicollis (MONBE). Последовательности также были выровнены при помощи MAFFT, однако теперь при попытке использовать модель TN93 возникала ошибка, связанная с нарушением правила треугольника при определнии длин ветвей. Вероятно, это связано с тем, что после удаления колонок, содержащих гэпы, для построения дерева оставалась слишком малая часть выравнивания. Поэтому для построения дерева (Рис. 2) была использована двухпараметрическая модель Кимуры (K2P).
fastme -i 12S_outgroup_aligned.phy -r -dK -o 12S_outgroup_K.phy
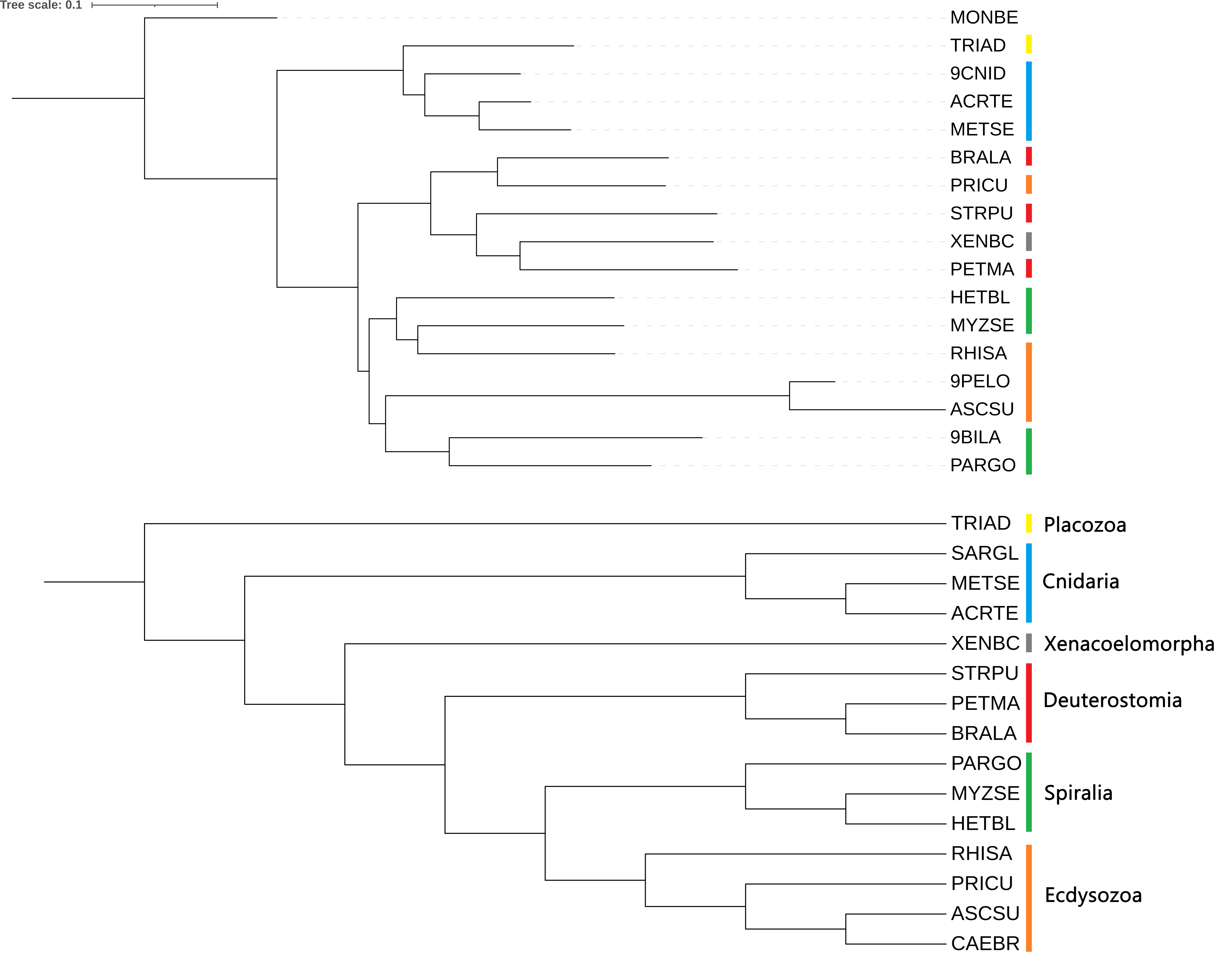
В данном случае ситуация изменилась не сильно. Однако теперь Trichoplax adhaerens образует монофилетическую группу с Cnidaria. Также можно заметить, что нематоды, как и всегда ранее, образуют отдельную группу.
Построение дерево с расчетом бутстреп-поддержки ветвей
Поскольку, как показали полученные деревья, реконструкция филогении достаточно далеких друг от друга групп Metazoa по последовательностям митохондриальной рРНК еще менее информативна, чем реконструкция по последовательностям цитохрома B, было принято решение построить дерево по цитохрому B с укоренением во внешнюю группу и расчетом бутстреп-поддержки ветвей.
В качестве внешней группы был выбран Saccharomyces cerevisiae (YEAST), так как белковая последовательность цитохрома B Monosiga brevicollis отсутствовала в Swiss-Prot. Последовательности были выровнены при помощи MAFFT и по выравниванию при помощи fastme с использованием метрики MtREV было построено дерево (Рис. 3).
fastme -i cyb_out_aligned.phy -r -pM -b 100 -o cyb_out_M.phy
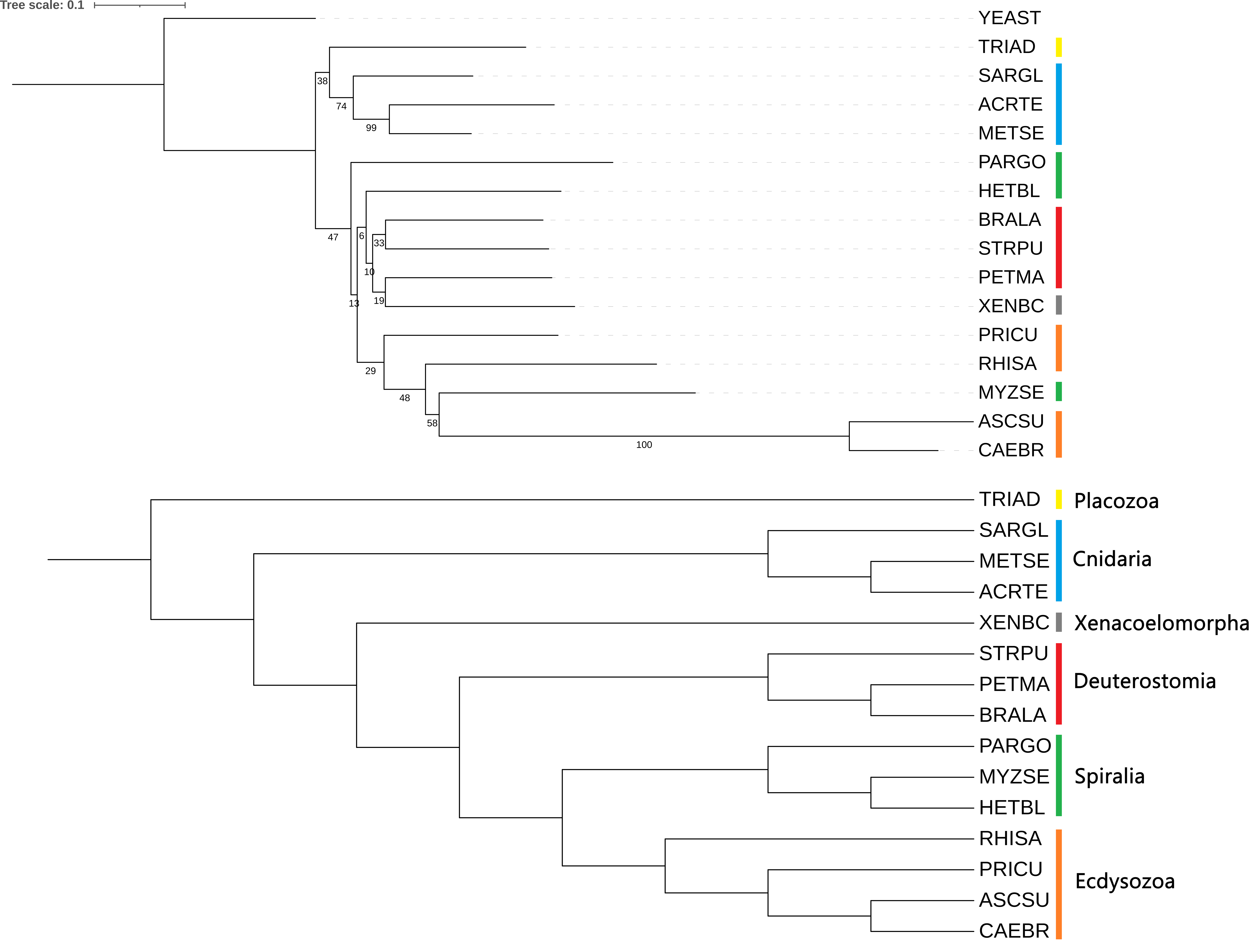
Как можно заметить, поддрежка почти всех ветвей оказалась низкой. Высокую поддержку имеют ветви отделяющие группу из двух нематод, а также группу из двух стрекающих из Hexacoralia (ACRTE и METSE). Также относительно высокую поддержку имеет ветвь, объединяющая всех стрекающих. Стоит заметить, что только эти ветви в точности совпадают с ветвями дерева, построенного по таксономии.
Таким образом, можно предположить, что высокую поддержку скорее будут иметь ветви, отражающие действительность и наоборот, однако это соотвествие не будет всегда однозначным.