Множественное выравнивание
Для своего множественного выравнивания я отобрала 33 гомолога моего белка YWHB_BACSU из других филумов бактерий, всего гомологов было 256, параметры поиска представлены в таблице 1, организмы, из которых взяты белки - в таблице 2. Филогения представлена на рисунке 1 и чуть более понятно в списке под картинкой.
|
Поиск |
Алгоритм BLAST |
Название базы данных |
Ограничения по таксонам |
Порог e-value |
Максимальное количество хитов |
|
По прокариотам |
BLASTP |
Refseq |
Firmicutes -Eukaryota |
1e-10 |
256 |
|
По эукариотам |
BLASTP |
Refseq |
Eukaryota |
10 |
4 |
Таблица 1. Параметры поиска гомологов с помощью BLAST.
|
Домен |
Филум/царство |
Название организма |
Количество белков |
|
Archaea |
Euryarchaeotes |
Methanosphaerula_palustris_E1-9c |
3 |
|
Methanoculleus_marisnigri_JR1 |
|||
|
Cenarchaeales |
Cenarchaeum_symbiosum_A |
1 |
|
|
Bacteria |
Proteobacteria |
Burkholderia_sp._CCGE1002 |
209 |
|
Ralstonia_solanacearum_IPO1609 |
|||
|
Cupriavidus_necator_N-1 |
|||
|
Cupriavidus_necator_N-1 |
|||
|
Azoarcus_sp._BH72 |
|||
|
Dechloromonas_aromatica_RCB |
|||
|
Pseudomonas_putida |
|||
|
Pseudomonas_putida |
|||
|
Agrobacterium_tumefaciens_str._Cherry_2E-2-2 |
|||
|
Micavibrio_aerunosavorus_EPB |
|||
|
Geobacter_metallireducens_GS-15 |
|||
|
Desulfobulbus_propionicus_DSM_2032 |
|||
|
Arcobacter_sp._L |
|||
|
Arcobacter_nitrofilis_DSM_7299 |
|||
|
Cyanobacteria |
Rivularia_sp._PCC_7116 |
13 |
|
|
Nostoc_sp._PCC_7524 |
|||
|
Leptolyngbya_sp._PCC_7375 |
|||
|
Coleofasciculus_chthonoplastes_PCC_7420 |
|||
|
Actinobacteria |
Amycolatopsis_decaplanina_DSM_44594 |
10 |
|
|
Amycolatopsis_azurea_DSM_43854 |
|||
|
Synergistales |
Thermovirga_lienii_DSM_17291 |
6 |
|
|
Synerstes_sp._3_1_syn1 |
|||
|
Thermales |
Thermus_scotoductus_SA-01 |
5 |
|
|
Thermus_oshimai_JL-2 |
|||
|
Aquificales |
Persephonella_marina_EX-H1 |
2 |
|
|
Hydrogenivirga_sp._128-5-R1-1 |
|||
|
Deferribacterales |
Flexistipes_sinusarabici_DSM_4947 |
2 |
|
|
Denitrovibrio_acetiphilus_DSM_12809 |
|||
|
Fusobacteria |
Sebaldella_termitidis_ATCC_33386 |
1 |
|
|
Holophagales |
Holophaga_foetida_DSM_6591 |
1 |
|
|
Firmicutes |
Bacillus_subtilis_st._168 |
|
|
|
Eukaryotes |
Animals |
Caenorhabditis elegans |
2 |
|
Culex quinquefasciatus |
|||
|
Green plants |
Arabidopsis thaliana |
1 |
|
|
Fungi |
Schizophyllum commune H4-8 |
1 |
Таблица 2. Встречаемость гомологов моего белка в различных таксонах.
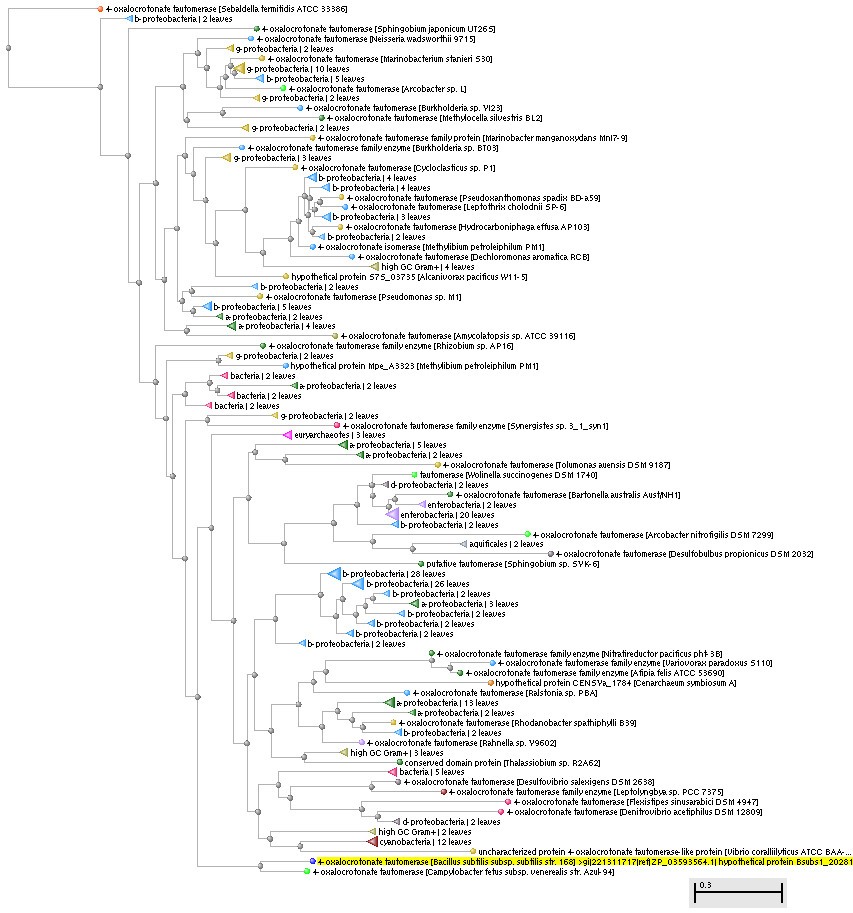
Рис. 1. Филогенетическое дерево гомологов моего белка среди прокариот.
Краткая филогения организмов, из которых были взяты гомологи белка YWHB_BASCU.
- Bacteria (251)
- Proteobacteria (211)
- b-proteobacteria (104)
- Burkholderiales (97)
- more... (7)
- g-proteobacteria (56)
- a-proteobacteria (41)
- d-proteobacteria (6)
- e-proteobacteria (4)
- Cyanobacteria (13)
- Nostocales (9)
- more... (4)
- Actinobacteria (10)
- Synergistales (6)
- Thermales (5)
- Aquificales (2)
- Deferribacterales (2)
- Fusobacteria (1)
- Holophagales (1)
- Archaea (4)
- Euryarchaeotes (3)
- Cenarchaeales (1)
- Eukaryotes (4)
- Animals (2)
- Arthropods (1)
- Nematodes (1)
- Green plants (1)
- Fungi (1)
Для множественного выравнивания использовала программу Muscle в интерфейсе JalView. Цветовая схема (таблица 3) собственная, аминокислоты раскрашены по свойствам. Выравнивание, сделанное программой выглядит так: рисунок 2. Сделаны следующие подписи: SECONDARY - вторичная структура белка - альфа-спирали и бета-тяжи, LIGAND - остатки, связанные с лигандом и BLOCKS - участки, выровненные по всей толщине. Яркость аминокислотных остатков определяется порогом консервативности 21.
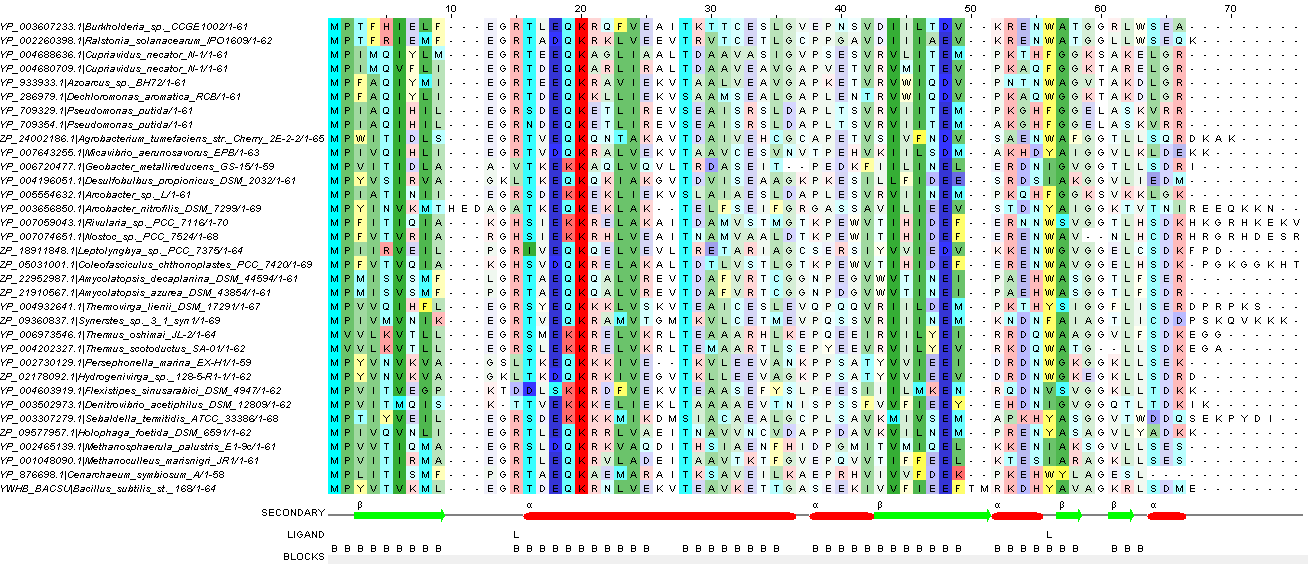
Рис. 2. Множественное выравнивание, сделанное программой Muscle. При окраске использовался порог консервативности 21 и собственная цветовая схема (таблица 3).
| Свойства аминокислотных остатков | Цвет |
| Алифатические | Зеленый |
| Ароматические | Желтый |
| Нейтрально заряженные | Голубой |
| Отрицательно заряженные | Синий |
| Положительно заряженные | Красный |
Таблица 3. Схема собственной окраски множественного выравнивания.
Далее провела редактирование выравнивания, опираясь на вторичную структуру моего белка. Что получилось, можно увидеть на рисунке 4, как это получилось, можно посмотреть по ссылке.
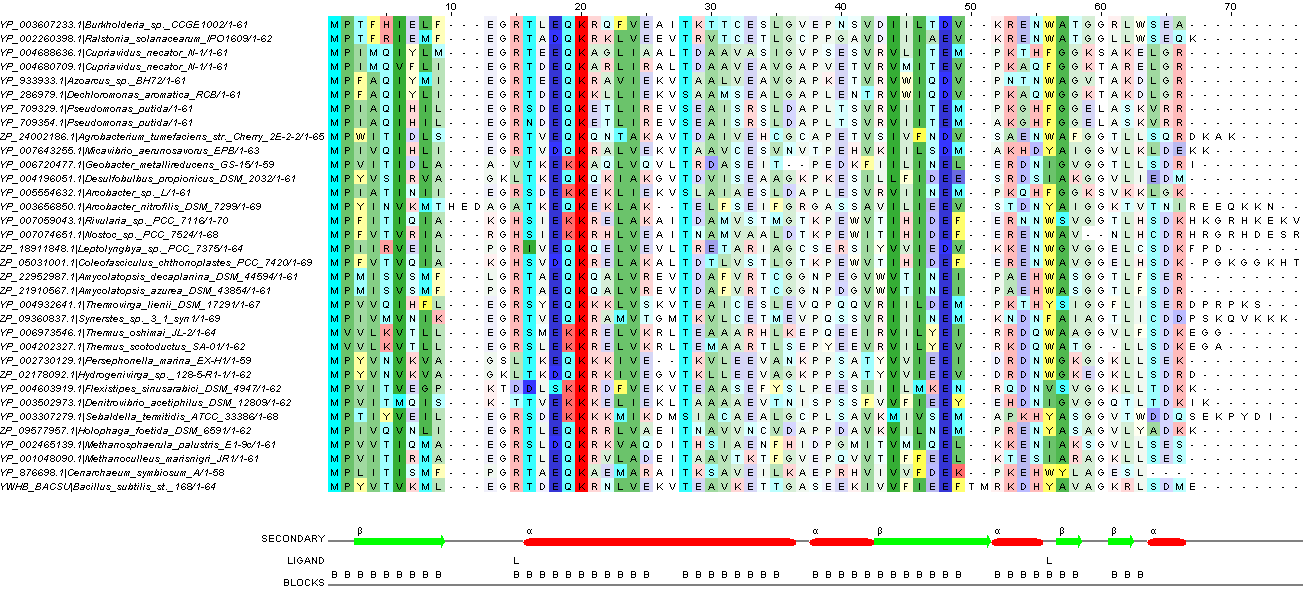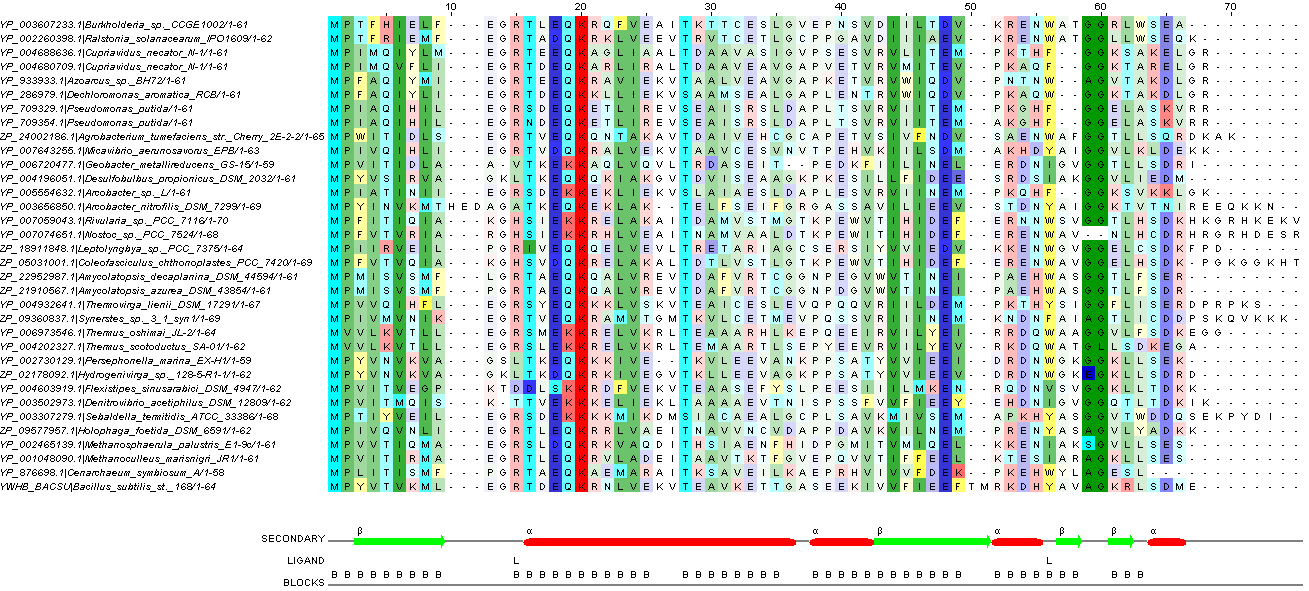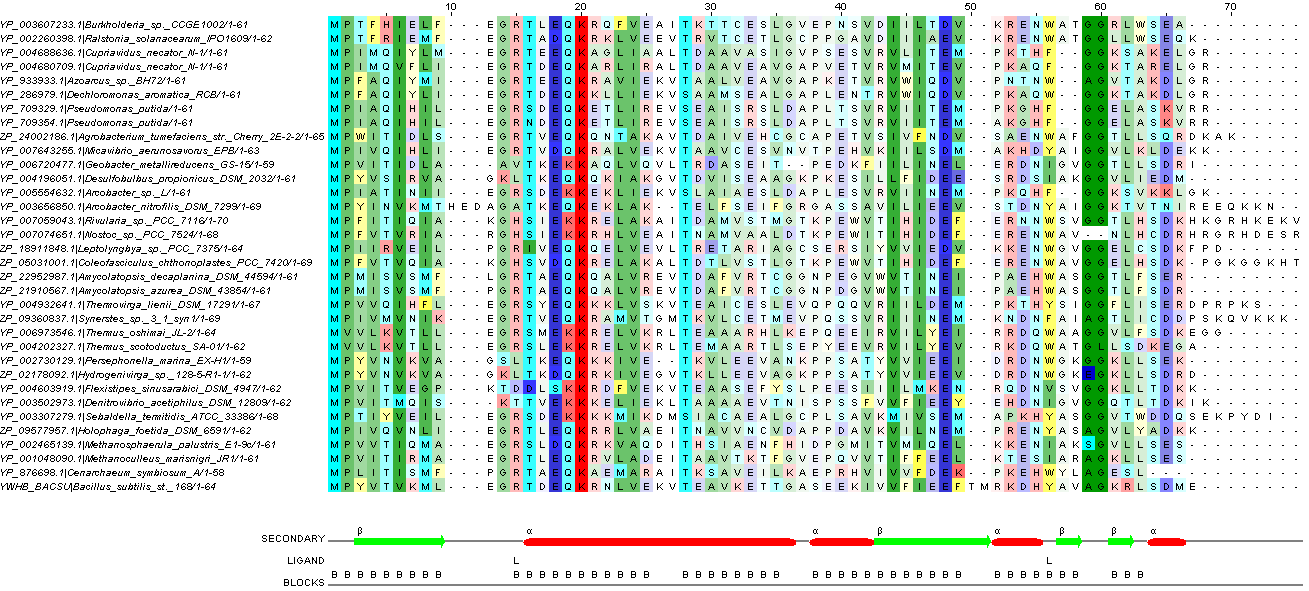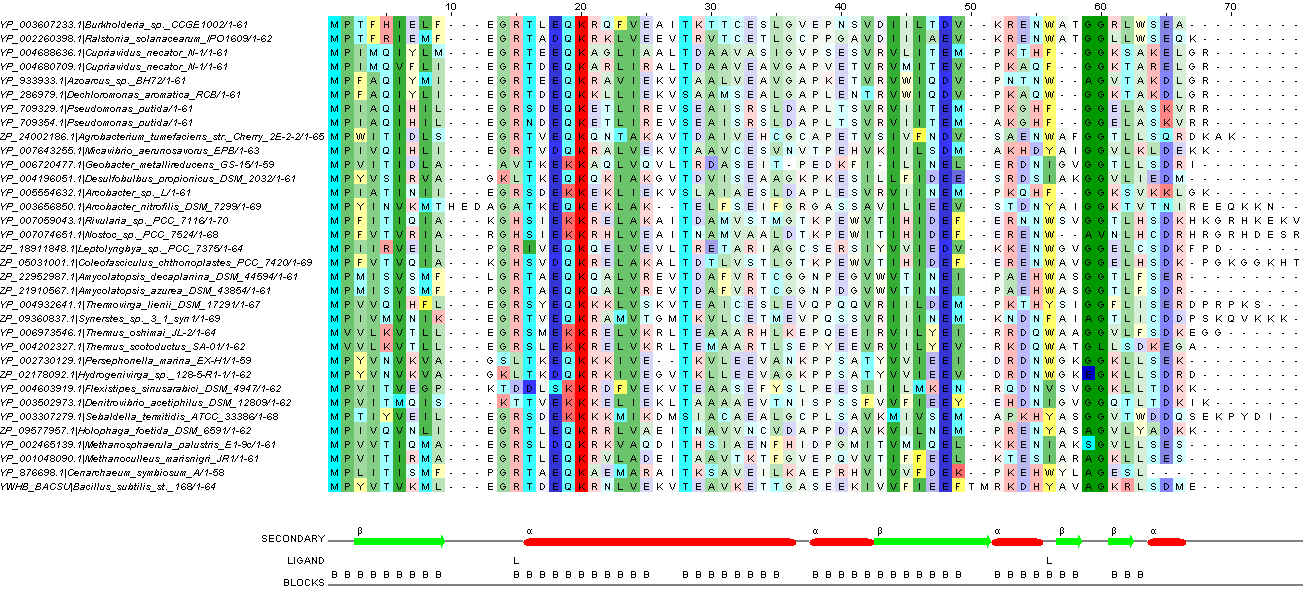{kind=link}
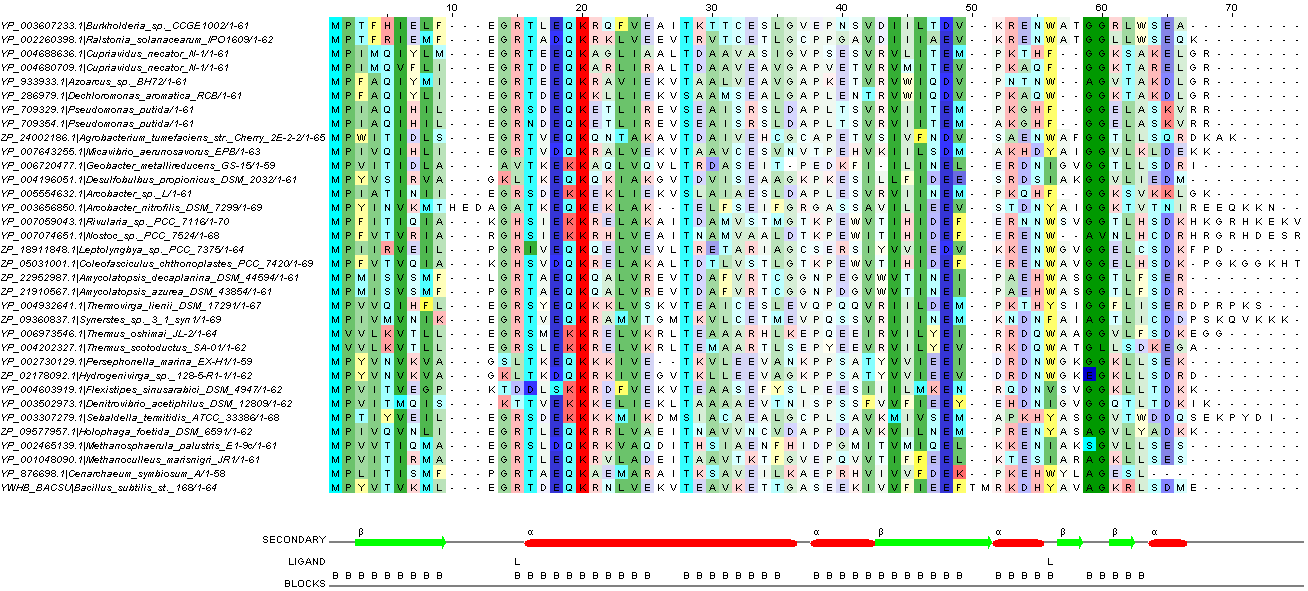
Рис. 3. Отредактированное множественное выравнивание. При окраске использовался порог консервативности 21 и собственная цветовая схема (таблица 3). Более яркие цвета означают большую консервативность.
Сделала ассоциацию трехмерной структуры своего белка с множественным выравниванием (рисунок 4). Цепь А (62 аминокислотных остатка) окрасилась в соответствии с выравниванием. Наиболее светлые участки наименее консервативны.
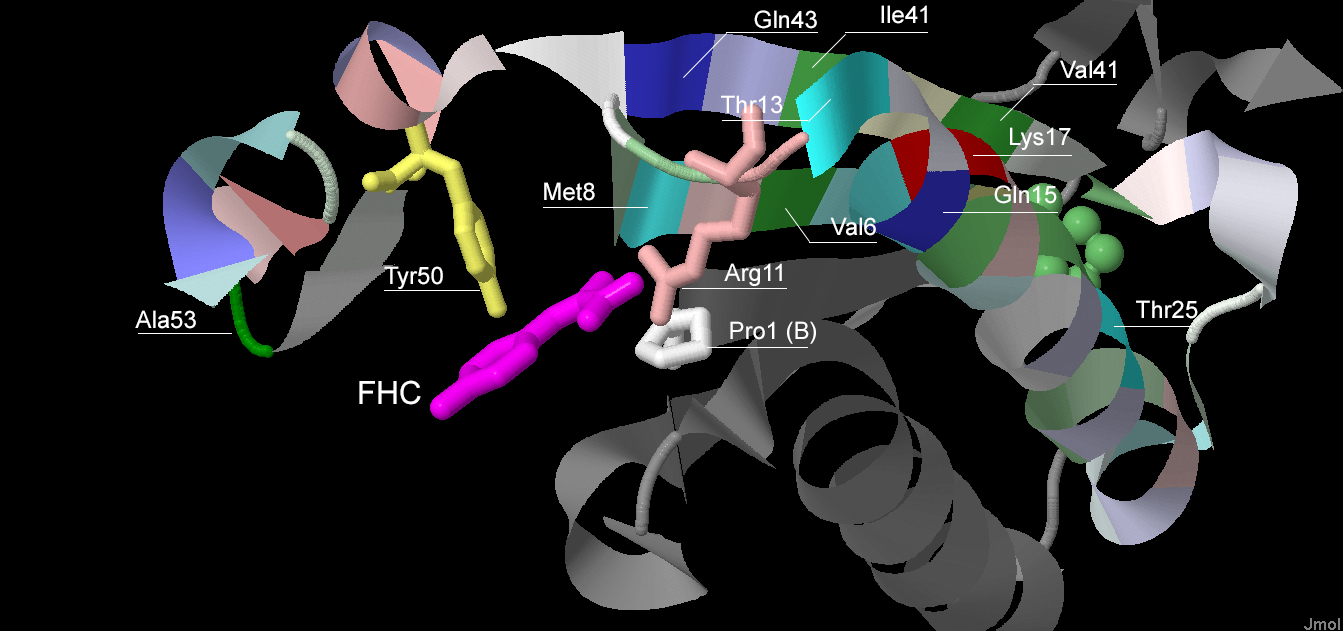
Рис. 4. Трехмерная структура белка YWHB, окрашенная в соответствии с консервативностью аминокислотных остатков. Яркая малиновая структура в центре - лиганд FHC. Подписаны консервативные остатки и остатки, связанные с лигандом. Связи с лигандом можно посмотреть здесь.
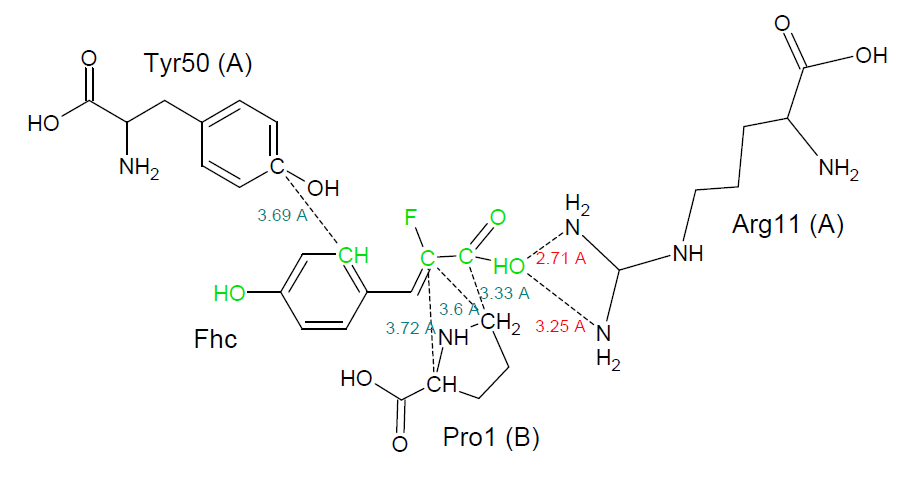{kind=link}
Достоверных гомологов для моего белка среди эукариот не нашлось. Я взяла 4 "гомолога" со значениями E-value от 1 до 10. Организмы представлены в таблице 1. Интереса ради попробовала сделать множественное выравнивание пяти последовательностей. Для этого увеличила последовательность свеого белка вдвое, будто бы это димер, как он есть в четвертичной структуре. Выравнивание можно увидеть на рисунке 5.
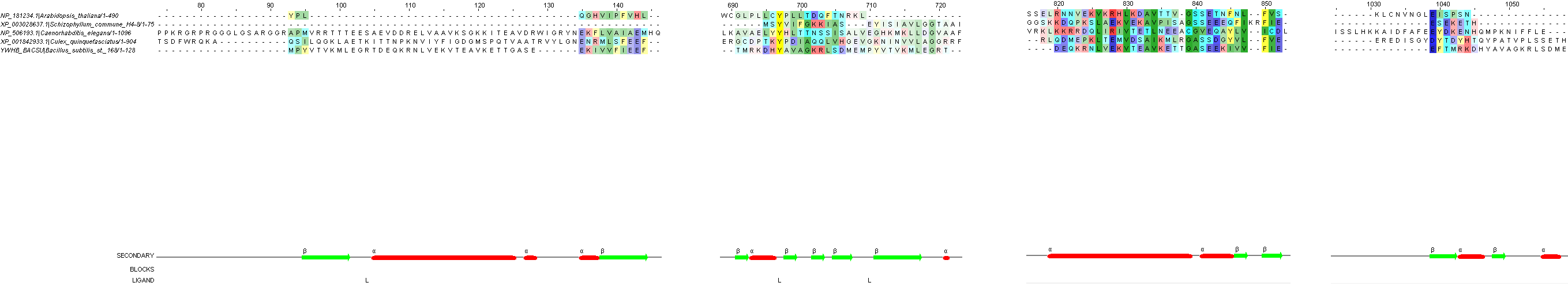
Рис. 5. Множественное выравнивание последовательностей из эукариот и моего белка. При окраске использовался порог консервативности 17 и собственная цветовая схема (таблица 3). Более яркие цвета означают большую консервативность.
Обсуждение
Проведя множественное выравнивание, мы убеждаемся, что большая консервативность соответствует важным участкам вторичной и третичной структуры. К примеру, заряженные аминокислоты консервативня, потому что образуют солевые мостики в альфа-спиралях, а маленькие алифатические аминокислоты обеспечивают поворот в участках петель. Важную функцию выполняют остатки, связывающие лиганд, они тоже довольно консервативны - судя по выравниванию, всего десять белков не могут в целом связывать FHC. У эукариот такой зависимости не обнаружено - белки слишком разные по структуре. Но, что удивительно, абсолютно консервативным остался триптофан и небольшие алифатические аминокислоты. Вероятно, это связано с необходимостью создавать выгодную конформацию молекулы.