практикум №3
методы
Укоренение. Сравнение деревьев. Бутстрэп
В данном практикуме построенные раннее дерево видов (таксономическое) и филогенетическая реконструкция по последовательностям белков будут сравниваться с реконструкцией дерева по нуклеотидным последовательностям.
Реконструкция дерева по нуклеотидным последовательностям.
Дерево реконструировано по генам малой РНК митохондриальных рибосом (12S rRNA), последовательности которых вырезаются из полного митохондриального генома вида. Для некоторых видов в базе данных ENA не нашлись последовательности полного генома. Вместо них были взяты близкородственные виды:
- Bos sauveli (BOSSA) заменен Bos javanicus (BOSJA)
замена в пределах рода; - Cephalorhynchus commersonii (CEPCM) заменен Cephalorhynchus heavisidii (CEPHA)
замена в пределах рода; - Cephalorhynchus eutropia (CEPEU) заменен Delphinus capensis (DELCA)
замена в пределах семейства, не изменяющая положение вида на дереве видов.
Филогенетическая реконструкция проведена на основе выравнивания последовательностей программой muscle с последующим построением дерева программой FastME (модель p-distance). На рис. 1 данное дерево сравнивается с деревом видов, на рис. 2 - с реконструированным по последовательностям белков деревом, построенным аналогичной программой и моделью.


По нуклеотидным последовательностям ошибочно не реконструирована только одна клада (та же, что и по белковым последовательностям). В реконструированном дереве исходно (то есть в дереве видов) внешний по отношению к кладе (MEGNO,BALMU) вид ESCRO становится сестринским виду MEGNO и, таким образом, образуется клада (BALMU,(ESCRO,MEGNO)).

Как отмечалось выше, деревья, реконструированные аналогичными програмамми FastME и моделью p-distance по нуклеотидным последовательностям 12S rRNA и белковым последовательностям цитохромов В (различающиея параметрами -d и -p соответственно), одинаковы (за исключением оценки расстояний и замены некоторых видов).
Укоренение во внешнюю группу.
Для укоренения во внешнюю группу был выбран представитель отряда Непарнокопытных - лошадь Equus caballus (HORSE). Последовательность 12S rRNA этого вида была добавлена в выравнивание нуклеотидных последовательностей, дерево реконструировано программой FastME (модель p-distance) и укоренено в ветвь, ведущую к внешней группе (рис.3).

Добавление внешней группы для укоренения изменило топологию дерева: (1) клада бегемотов (HIPAM,HEXLI) стала сестринской по отношению к полорогим (PELCP,(BOVIN,BOSJA)), в то время как на дереве видов последнюю кладу можно называть внешней. Также интересно, что ошибочно не реконструированная на всех деревьях клада (ESCRO,(MEGNO,BALMU)) в данном случае имеет другое строение: внешним к двум сестринским видам становится MEGNO, а виды ESCRO и BALMU объединяются в одну кладу (2). При этом, что интересно, если взять более эволюционно далекий вид для укоренения (например, амурского ежа Erinaceus amurensis, 9EUTH - представителя отряда Насекомоядных), то изменение топологии (1) не наблюдается, при этом сохраняется изменение (2) (рис. 4).

Бутстреп.
Чтобы проверить, действительно ли неправильно реконструированные ветви имеют более низкую поддержку, использовано дерево с наибольшим количеством ошибочно не реконструированных клад - реконструированное по нуклеотидным последовательностям и укорененное в ветвь, ведущую к внешней группе HORSE (рис. 3). При построении дерева использована бутстреп-поддержка (дополнительный параметр -b 100) для создания программой 100 реплик (рис. 5).
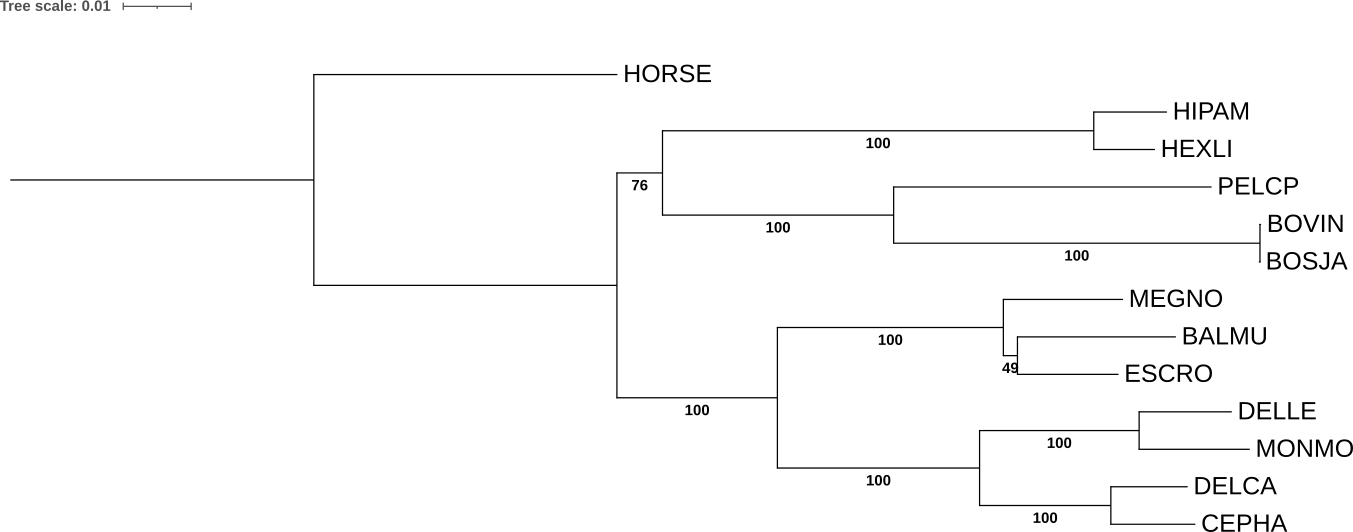
Клады, которые не претерпевали изменения при различных реконструкциях, имеют максимальное значение 100. Обусловленное укоренением в ветвь, ведущей к данной внешней группе, изменение топологии (1), то есть клада с сестринскими бегемотами и полорогими (см. прошлый пункт), имеет значение 76 - это умеренно высокая поддержка, свидетельствующая, вероятно, о статистической значимости ветвления, хоть это и не совпадает с деревом видов. Самый спорный узел усатых китов, исходно ((BALMU,MEGNO),ESCRO), который реконструируется по-разному в зависимости от наличия укоренения всего дерева в ветвь внешней группы, имеет поддержку 49, то есть реконструирован некорректно.