Занятие 4: Комплексы ДНК-белок
Поиск ДНК-белковых контактов в заданной структуре
В программе RasMol с помощью команды define можно задавать множества атомов. Определим множества следующих атомов:
- множество атомов кислорода 2'-дезоксирибозы (set1);
- множество атомов кислорода в остатке фосфорной кислоты (set2);
- множество атомов азота в азотистых основаниях(set3);
Создадим скрипт, вызов которого даёт в программе RasMol последовательно следующие изображения:
- вся структура,
- только ДНК (в проволочной модели);
- проволочная модель ДНК с выделенными шариками множества атомов set1,
- проволочная модель ДНК с выделенными шариками множества атомов set2,
- проволочная модель ДНК с выделенными шариками множества атомов set3.
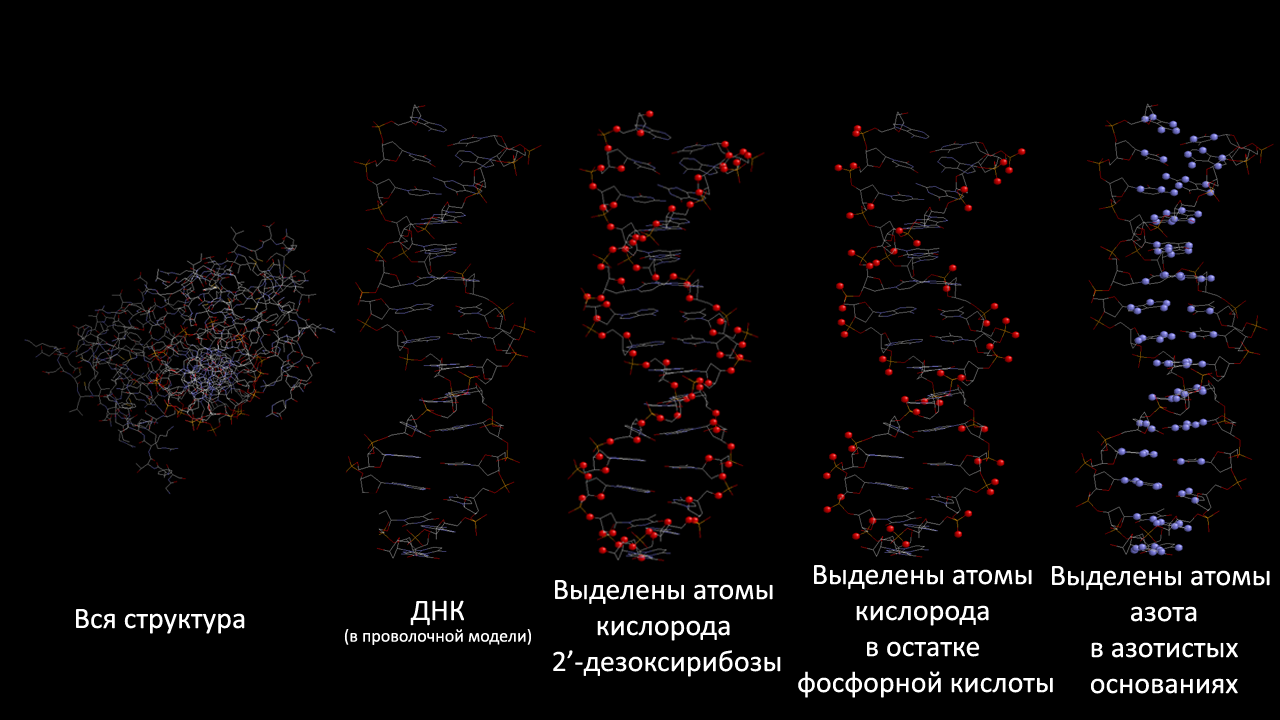
Скрипт представлен в файле dnascript.spt.
Описание ДНК-белковых контактов в заданной структуре
Для исследования ДНК-белковых контактов использована структура 1by4.pdb.
Полярными будем сичтать атомы кислорода и азота, а неполярными - атомы углерода, фосфора и серы.
Полярным контактом назовем ситуацию, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5Â. Аналогично, неполярным контактом будем считать пару неполярных атомов на расстоянии меньше 4.5Â.
Написан скрипт dna_protein_contacts.spt, последовательно выводящий на экран изображения структуры и атомов, необходимых для определения ДНК-белкового контакта соответствующего типа. Результаты определения числа контактов приведены в следующей таблице:
Контакты разного типа в комплексе 1by4.pdb
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 5 | 17 | 22 |
| остатками фосфорной кислоты | 23 | 18 | 41 |
| остатками азотистых оснований со стороны большой бороздки | 4 | 8 | 12 |
| остатками азотистых оснований со стороны малой бороздки | 0 | 2 | 2 |
Если ограничить исследование ДНК-белковых контактов только контактами ДНК с цепью B белка, то получим следующие значения для количества контактов разного типа:
Контакты разного типа в комплексе 1by4.pdb (рассмотрена только цепь B белка)
| Контакты атомов цепи B белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 3 | 7 | 10 |
| остатками фосфорной кислоты | 15 | 12 | 27 |
| остатками азотистых оснований со стороны большой бороздки | 3 | 3 | 6 |
| остатками азотистых оснований со стороны малой бороздки | 0 | 1 | 1 |
Мы видим, что наибольшее число контактов (как полярных, так и неполярных) - между белком и остатками фосфорной кислоты. Это вполне объяснимо, ведь атомы остатков фосфорной кислоты наиболее доступны для взаимодействия благодаря своему расположению. Мы также наблюдаем отсутствие полярных контактов атомов белка с остатками азотистых оснований со стороны малой бороздки. Это, вероятно, является следствием пространственной недоступности атомов азотистых оснований, направленных в сторону малой бороздки.
В качестве примера ниже приведены изображения, полученные в программе RasMol при помощи скрипта dna_protein_contacts.spt при изучении контактов атомов белка с остатками азотистых оснований со стороны большой бороздки.

Получение популярнуой схемы ДНК-белковых контактов
Для получения популярной схемы ДНК-белковых контактов использована программа nucplot. Так как данная программа работает только со старым форматом .pdb, для работы с ней был использован файл 1by4_old.pdb, полученный с помощью программы remediator:
nucplot 1by4_old.pdb
Одним из полученных файлов является файл nucplot.ps, содержание которого представлено на изображении ниже:
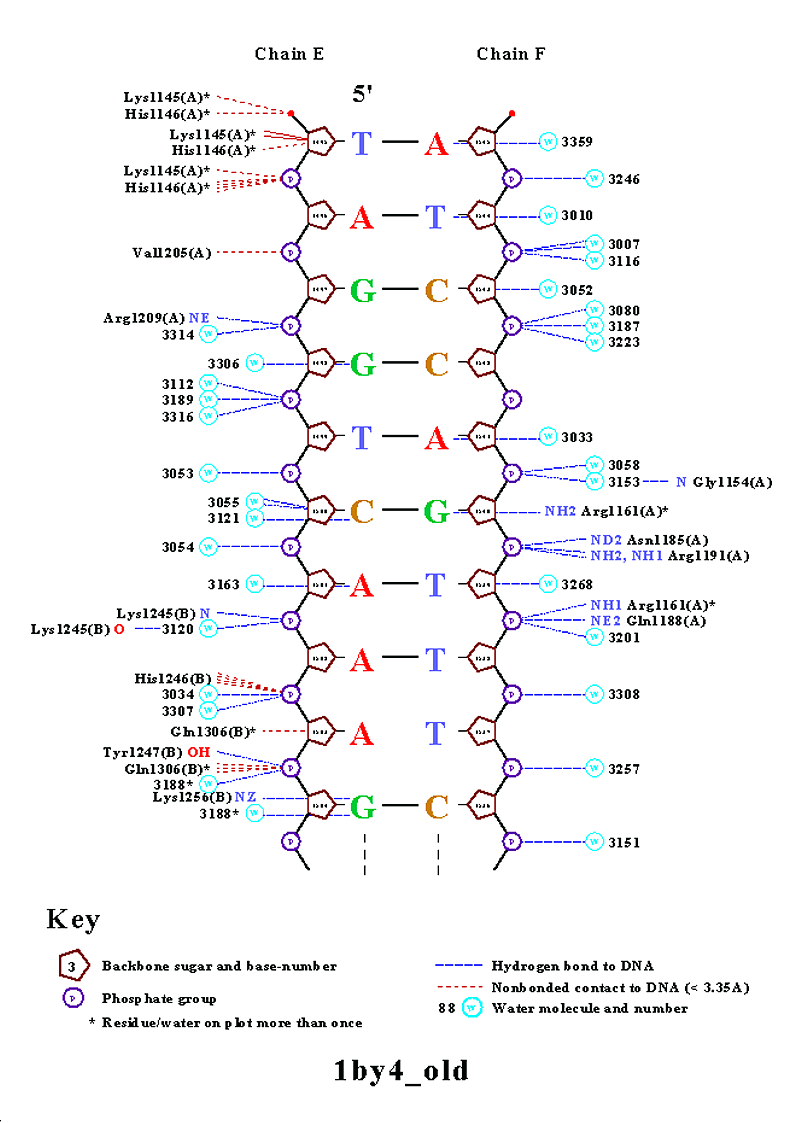
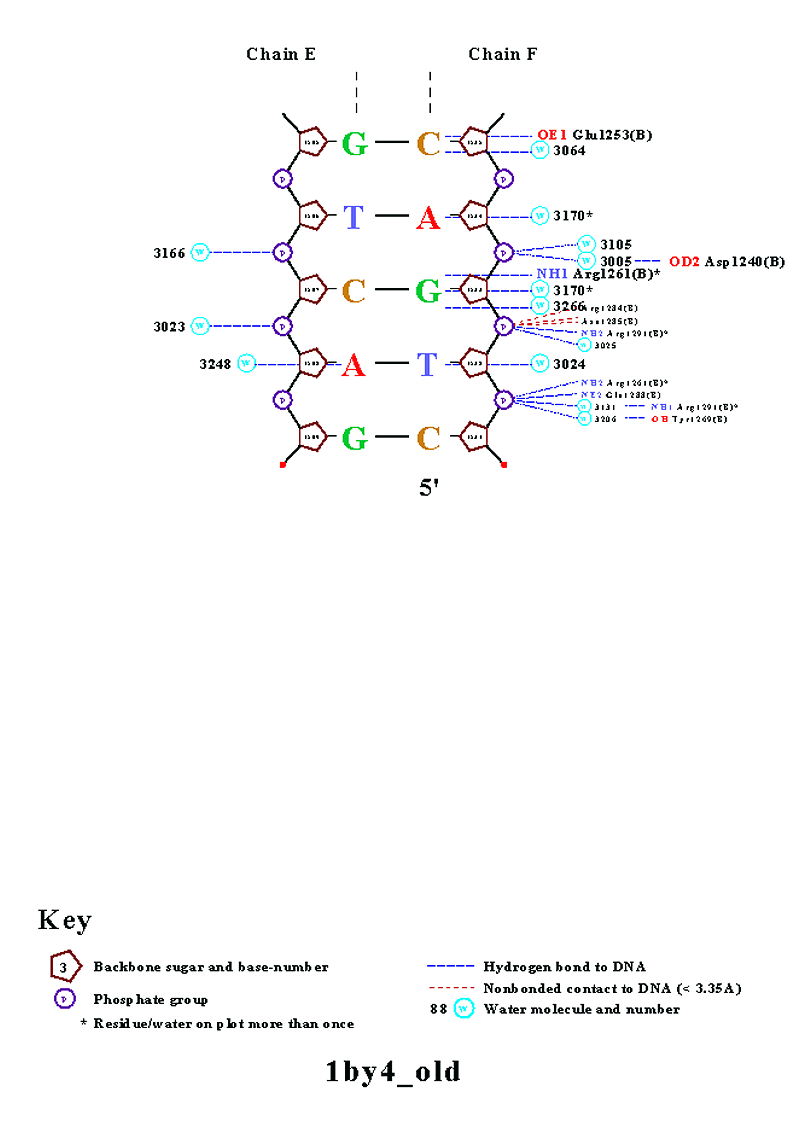
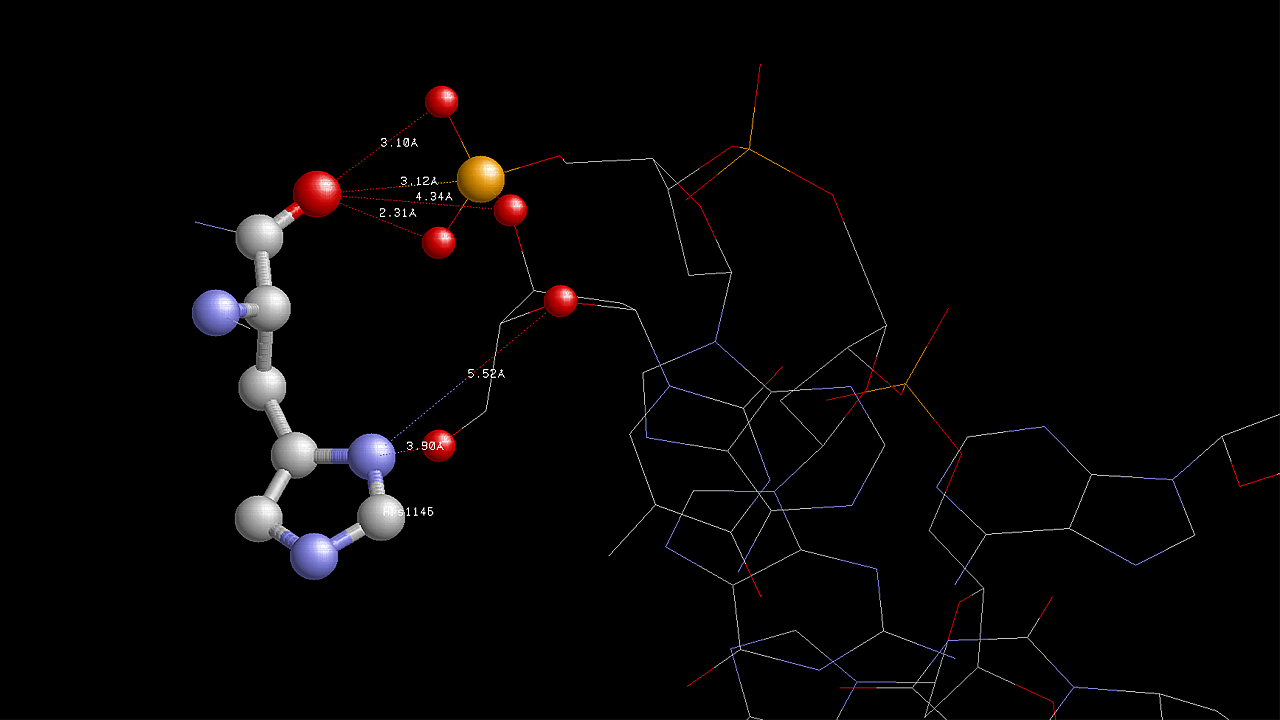
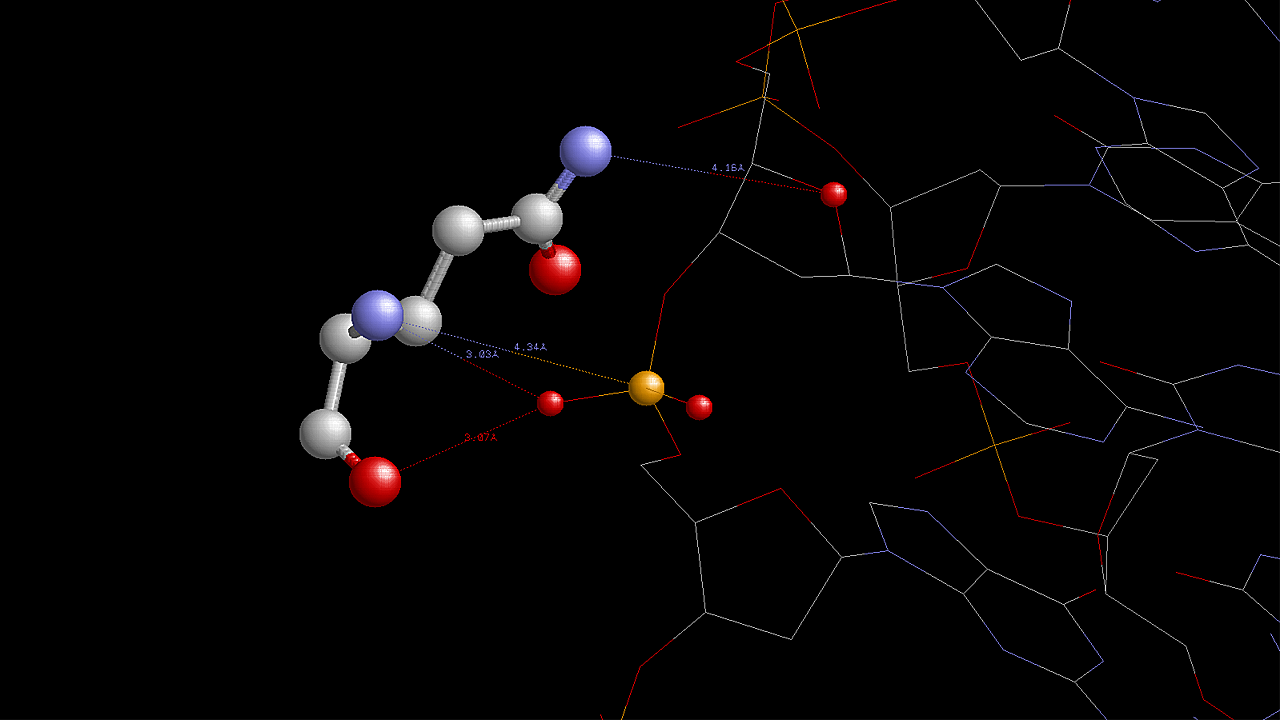
На данной схеме видно, что наибольшее число контактов с ДНК образует аминокислотный остаток His1146(A) (6 контактов). Контакты этого аминокислотного остатка с ДНК показаны на следующем изображении, полученным с помощью программы RasMol:
Если в рассмотрение брать только цепь B белка, то наибольшее число контактов с ДНК образуют His1246(B) и Gln1306(B) (по 4 контакта). Изображение контактов последнего из перечисленных аминокислотных остатков с ДНК приведено ниже:
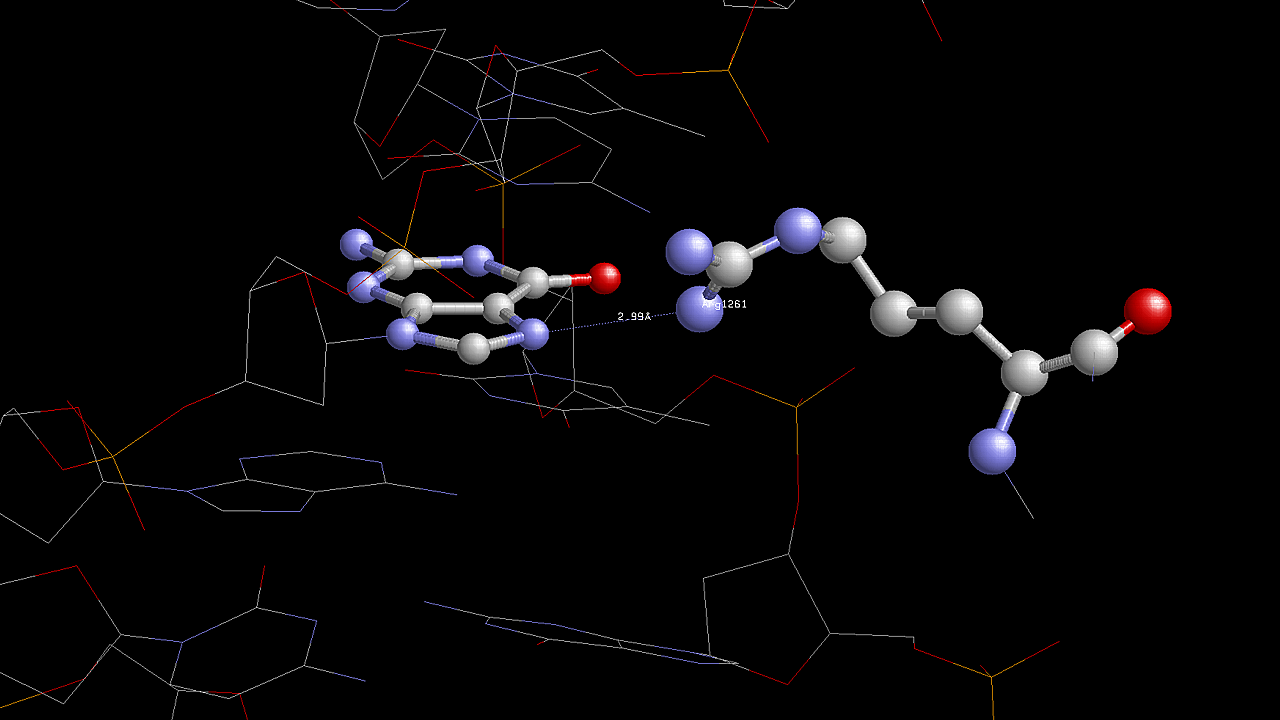
Для участия в распознавании последовательности ДНК аминокислотный остаток, очевидно, должен связываться с азотистым основанием. Как мы видим на схеме ДНК-белковых контактов, таких остатков немного. Это Lys1256(B), Arg1161(A), Glu1253(B) и Arg1261(B). Так как в этом списке аргинин является наиболее часто встречающимся азотистым основанием, ниже приведено изображение контактов аргинина, в частности Arg1261(B), с ДНК:
Предсказание вторичной структуры заданной тРНК
Предсказание вторичной структуры тРНК путем поиска инвертированных повторов
Программа einverted из пакета EMBOSS позволяет найти инвертированные участки в нуклеотидных последовательностях. Найдём возможные комплементарные участки в последовательности исследуемой тРНК (структуры 2xdi.pdb, последовательность тРНК можно найти в файле 2dxi_c.fasta) с помощью следующей команды:
einverted 2dxi_c.fasta.txt
При параметрах по умолчанию инвертированных участков найдено не было. Поэтому для наиболее удачного предсказания были подобраны следующие параметры:
- Gap penalty: 15
- Minimum score threshold: 25
- Match score: 10
- Mismatch score: -15
В результате мы получили файлы 2dxi_c.inv и 2dxi_c_einverted.fasta, информация из которых использована для заполнения таблицы ниже.
Предсказание вторичной структуры тРНК по алгоритму Зукера
Алгоритм Зукера реализует программа mfold. Нас будет также интересовать один из параметров этой программы - P, который указывает, на сколько процентов выдаваемое предсказание структуры может отличаться по своей вычисленной энергии от оптимального (соответственно, чем больше значение этого параметра, тем больше вариантов предсказания будет выдано).
Можно использовать программу mfold на сервере Mobyle @Pasteur.
Первое исполнение программы mfold для последовательности 2dxi_c.fasta с параметром P = 0% приводит к получению предсказания, очень близкого к реальной структуре. По мере увеличения значений параметра P (с шагом в 5%) количество вариантов предсказаний увеличивается. Однако предсказание, полученное при P = 0%, остаётся наиболее близким к реальной структуре. Изображение с лучшим предсказанием, а также результаты предсказания по алгоритму Зукера приведены ниже.
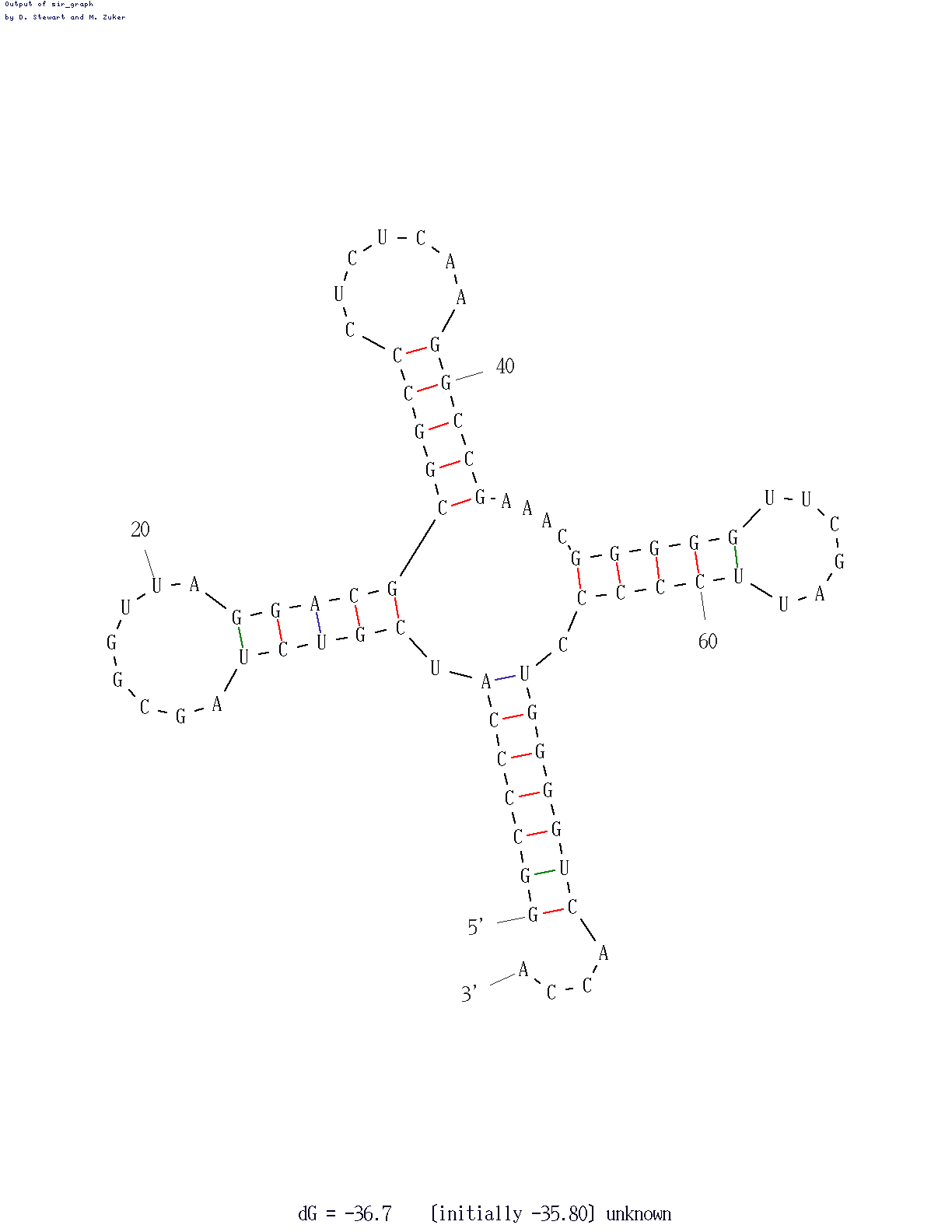
Реальная и предсказанная вторичная структура тРНК из файла 2dxi.pdb
| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
| Акцепторный стебель | 5'-501-572-3' 5'-502-571-3' 5'-503-570-3' 5'-504-569-3' 5'-505-568-3' 5'-506-567-3' 5'-507-566-3' Всего 7 пар |
предсказано 5 пар из 7 реальных | предсказано 7 пар из 7 реальных |
| D-стебель | 5'-510-522-3' 5'-511-523-3' 5'-512-524-3' 5'-513-525-3' Всего 4 пары |
предсказано 3 пары из 4 реальных | предсказано 4 пары из 4 реальных |
| T-стебель | 5'-549-565-3' 5'-550-564-3' 5'-551-563-3' 5'-552-562-3' 5'-553-561-3' Всего 5 пар |
предсказано 0 пар из 5 реальных | предсказано 4 пары из 5 реальных |
| Антикодоновый стебель | 5'-526-544-3' 5'-527-543-3' 5'-528-542-3' 5'-529-541-3' 5'-530-540-3' 5'-531-539-3' 5'-532-538-3' Всего 7 пар |
предсказано 5 пар из 7 реальных | предсказано 5 пар из 7 реальных |
| Общее число канонических пар нуклеотидов | 19 | предсказано 13 канонических пар | предсказано 18 канонических пар |
Ссылки
- Файл dnascript.spt.
- Файл dna_protein_contacts.spt.
- Файл dna_protein_contacts_B_chain.spt.
- Файл 2dxi.pdb.
- Файл 2dxi_c.fasta.
- Файл 2dxi_c.inv.
- Файл 2dxi_c_einverted.fasta.
- Файл 1by4.pdb.
- Файл 1by4_old.pdb.
- Файл nucplot.ps.