| Учебный сайт Саши Погорельской |

|
|
||||||
| Главная | Семестры | Скрипты | Обо мне | Ссылки | ||
|
|
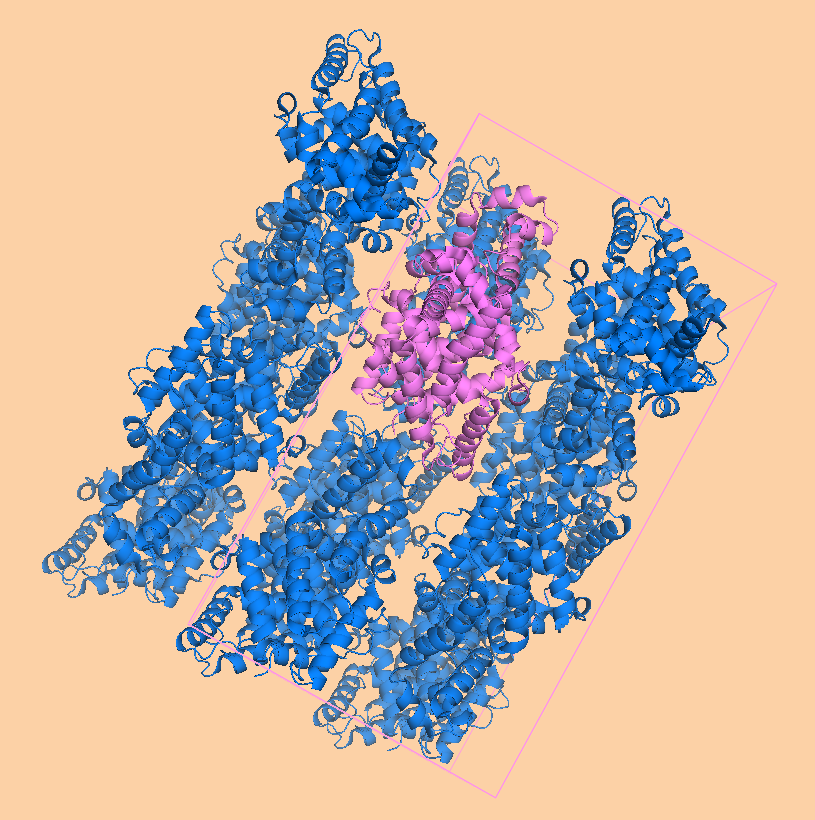
Восстановление кристалла из PDB файла Для работы использовался тот же белок, что и во множестве предыдущих практикумов: транскрипционный репрессор семейства TetR. Согласно PDB-файлу, кристаллографическая ячейка образована ортогональными векторами с длинами, равными 4.433, 110.721 и 126.030 ангстрем. Такая ячейка содержит 16 молекул белка, или 8 биологических единиц, так как репрессор работает в форме димера. Таким образом, нужно 7 движений, чтобы восстановить кристалл по асимметрической единице. 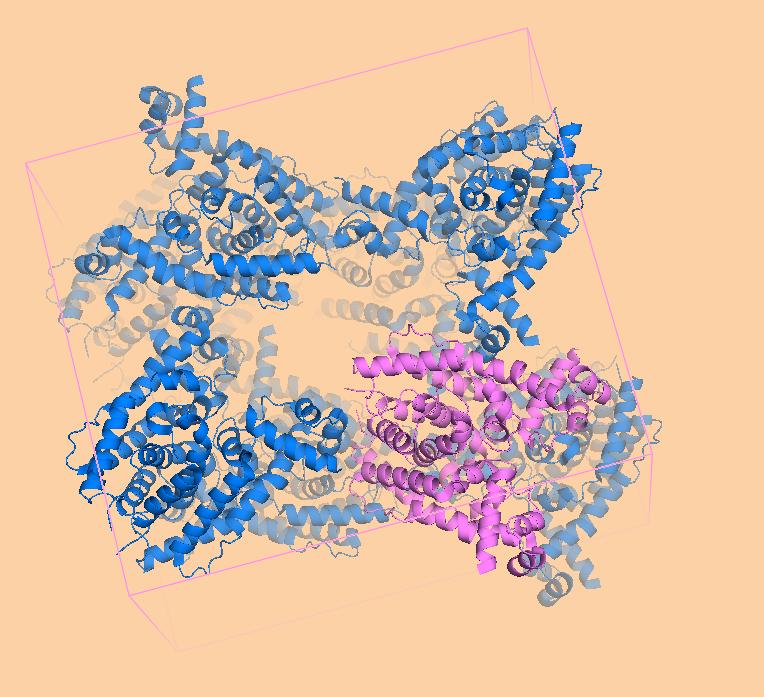 Рисунок 1. Расположение молекул в одном из слоев кристаллографической ячейки. Розовым цветом показан димер, описанный в PDB-файле, параллелепипед - 1 кристаллографическая ячейка. Изображение получено с помощью pymol Расположение молекул в одном из слоев кристаллографической ячейки показано на рисунке 1. Видно, что соседние димеры имею разную ориентацию относительно центра ячейки. Молекулы димеров не попадают полностью в одну ячейку, отдельные атомы принадлежат соседним. В кристаллографической ячейке можно выделить 2 слоя (рисунок 2) и 2 ряда (рисунок 3) по 4 димера.
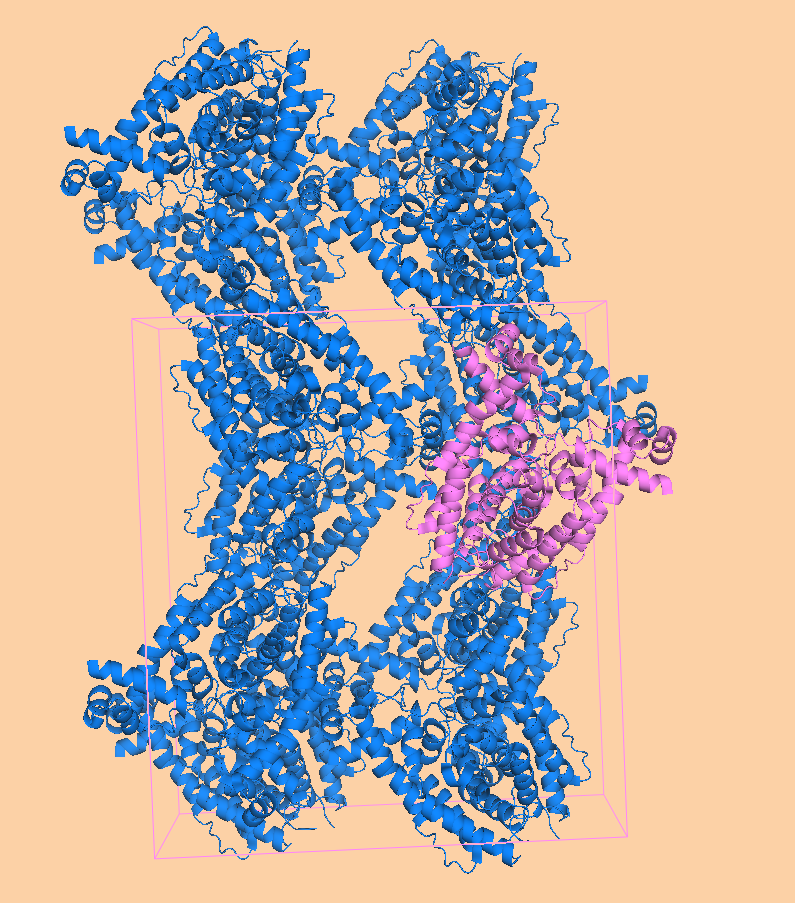
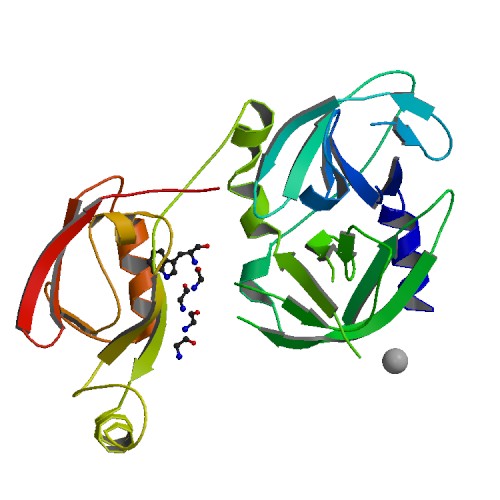
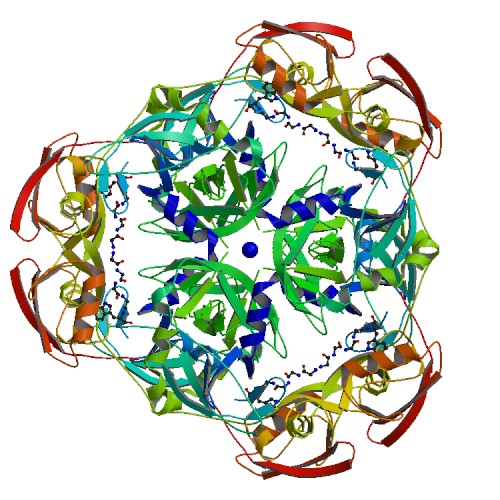
Каждая пара молекул (димер) имеет 6 возможных плоскостей контакта с другими элементами решетки. Из-за симметрии число различных плоскостей сокращается до 3. Предполагаемые области контактов указаны на рисунке 4. Расстояние между атомами, принадлежащих соседним слоям молекул (если смотреть в направлении стрелочки на рисунке 4), слишком велико, поэтому между слоями контактов нет (рисунок 5). 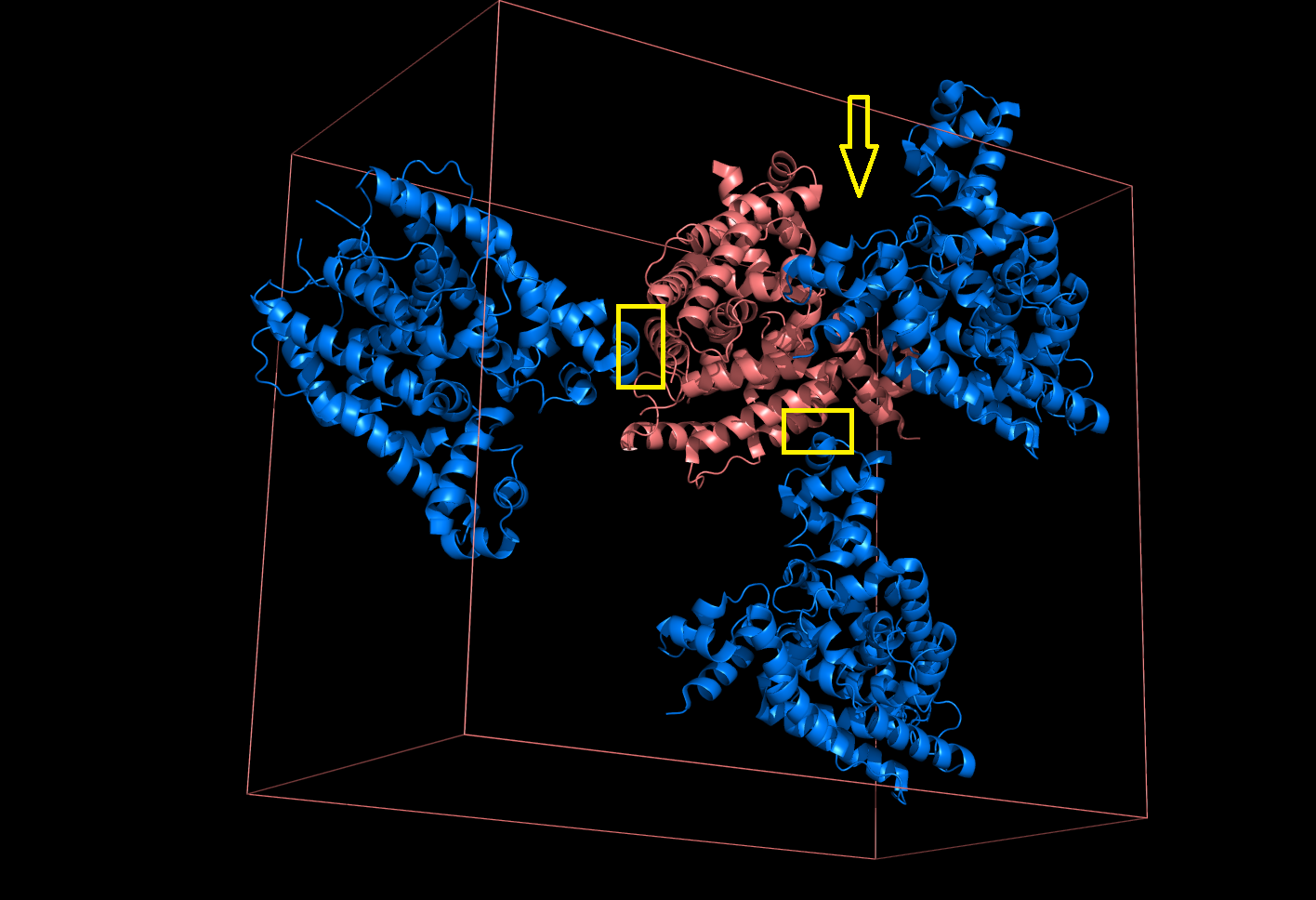 Рисунок 4. Предполагаемые области контакта димера с соседними молекулами в кристалле. Розовым цветом показан димер, описанный в PDB-файле, параллелепипед - 1 кристаллографическая ячейка. Изображение получено с помощью pymol 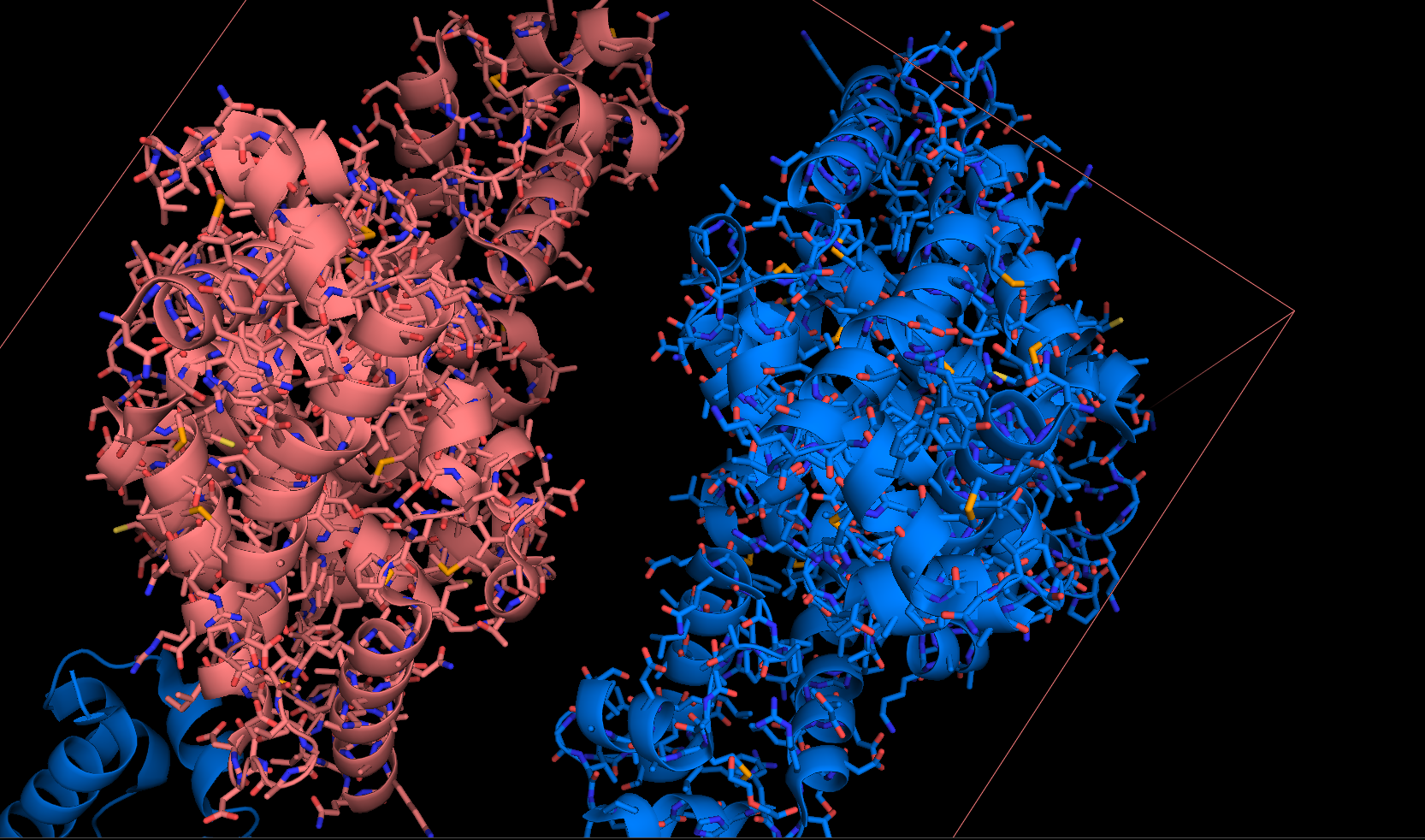 Рисунок 5. Соседние слои в кристаллографической ячейке (по направлению стрелочки на рисунке 4). Розовым цветом показан димер, описанный в PDB-файле, параллелепипед - 1 кристаллографическая ячейка. Изображение получено с помощью pymol Области, отмеченные на рисунке 4 прямоугольниками, содержат контакты между соседними молекулами. На рисунках 6 и 7 показаны атомы, находящиеся на расстоянии меньше 4 ангстрем. Для молекул соседних рядов (рисунок 6) можно отметить 2 солевых мостика с расстоянием между атомами 2.9 и 3.1 ангстрем. Также возможен контакт с длиной мостика 3.7 ангстрем. Атомы, принадлежащие молекулам одного ряда (рисунок 7), располагаются достаточно близко друг к другу, но довольно часто эти атомы являются атомами кислорода. Возможно, кислород остатка глутама и тирозина координируют один и тот же атом водорода, но это маловероятно для двух спиртовых групп (остатка тирозина и этандиола, который зажат между молекулами белка). Похоже, что наличие этих взаимодействий обусловлено лишь плотной упаковкой молекул в кристалл. А маленькие расстояния между атомами случайны, так как их довольно мало. В растворе энергии таких связей не хватит для ужержания белков вместе. 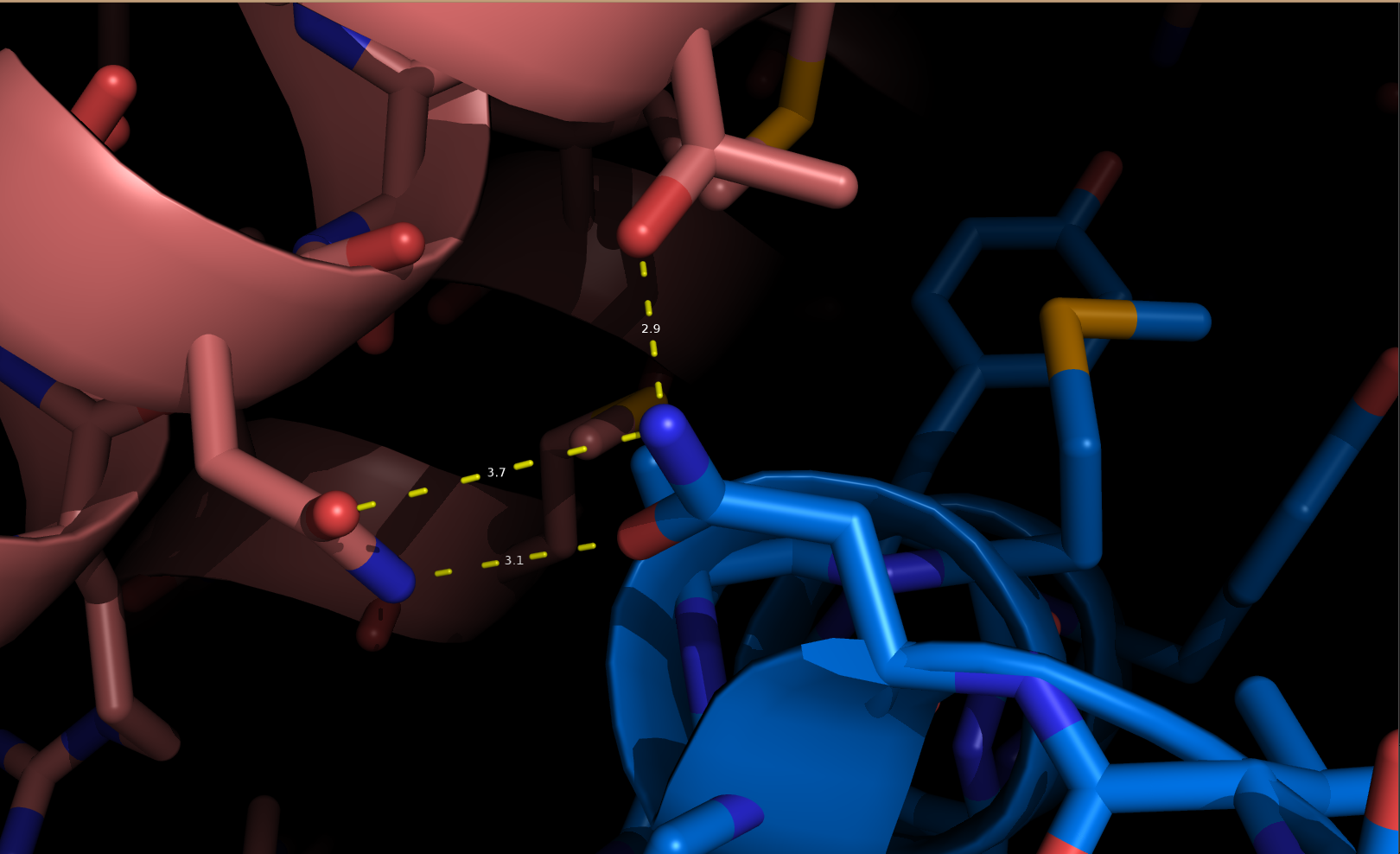 Рисунок 6. Участок между соседними рядами в кристаллографической ячейке (правый прямоугольник на рисунке 4). Розовым цветом показан димер, описанный в PDB-файле, параллелепипед - 1 кристаллографическая ячейка. Изображение получено с помощью pymol 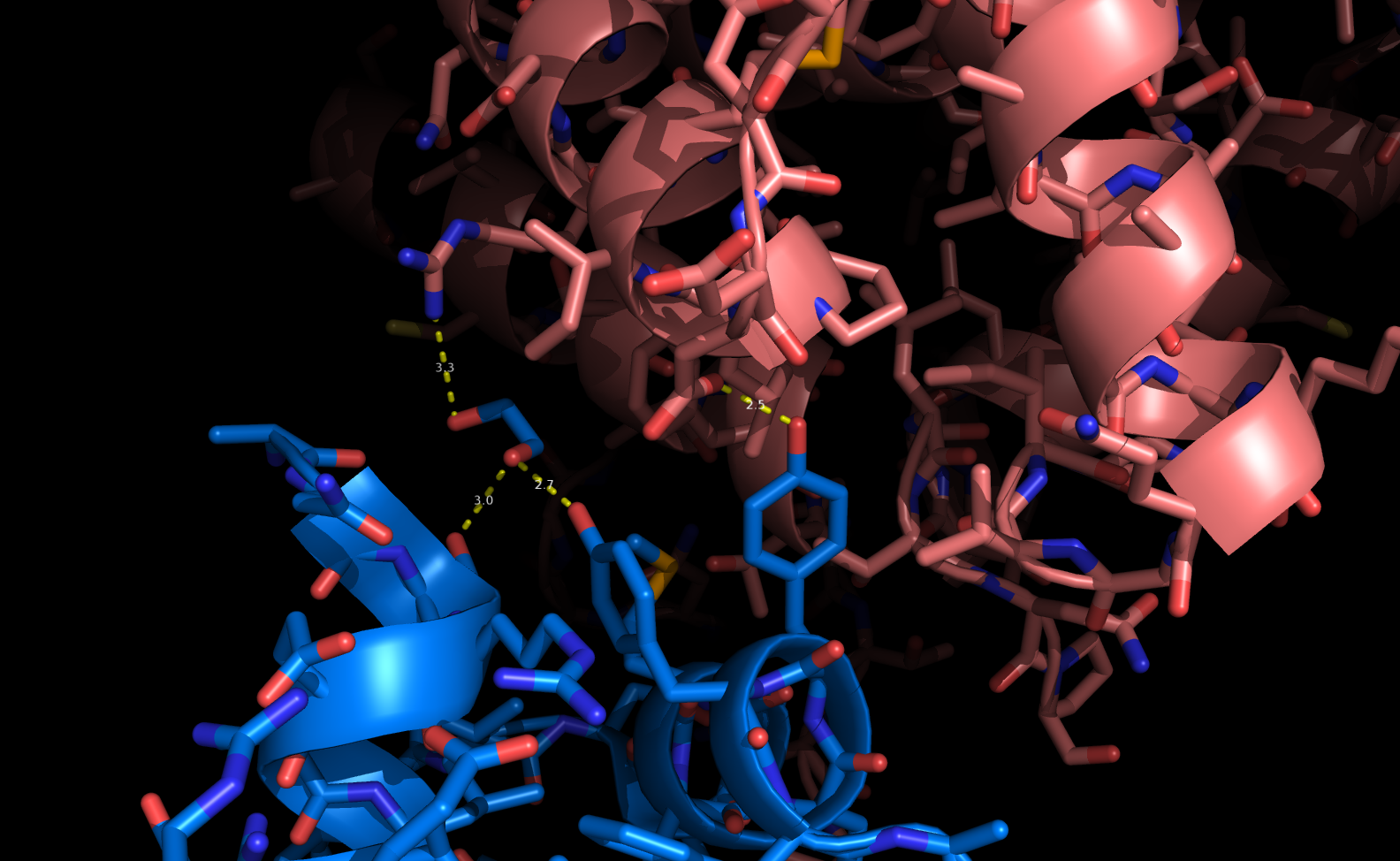 Рисунок 7. Участок между молекулами одного ряда в кристаллографической ячейке (левый прямоугольник на рисунке 4). Розовым цветом показан димер, описанный в PDB-файле, параллелепипед - 1 кристаллографическая ячейка. Изображение получено с помощью pymol Случай странного расположения белковых цепей В структуре ДНК-белкового комплекса с идентификатором 3hdd есть необычная область. На рисунке 8 видно, что один из гомеодоменов наполовину "висит" в воздухе - контактирует с ДНК только с одной стороны. Однако, если изобразить соседнюю кристаллографическую ячейку (рисунок 9) становится ясно, почему так происходит. Кусочки цепей ДНК, которые входят в кристалл, имеют так называемые липкие концы: выступающий 5' аденин и на другом конце 3' тимин. При кристаллизации они образуют пару, как это заметно из рисунка 9, и именно в этой бороздке, образованной цепями соседних ячеек, связана молекула белка. 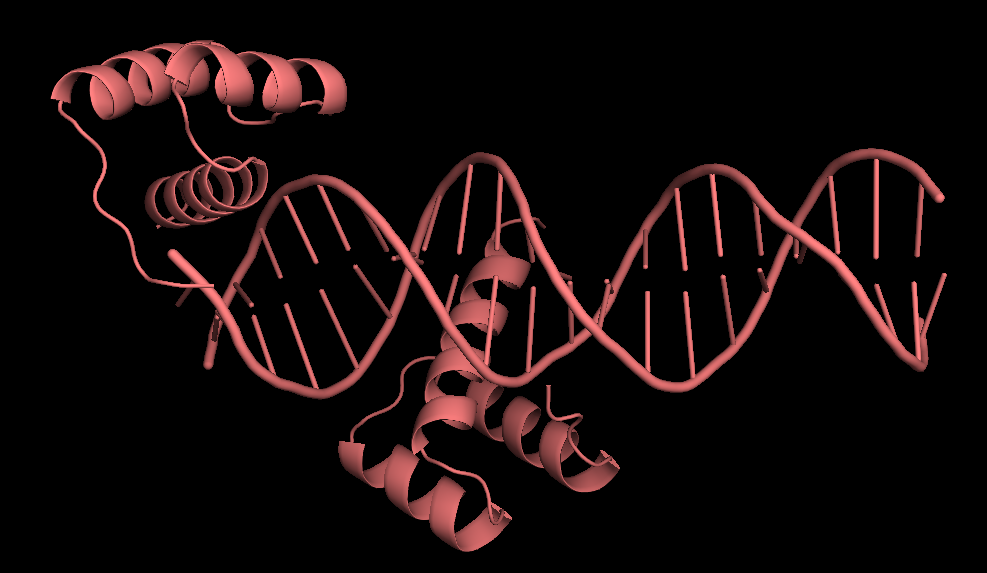 Рисунок 8. Структура ДНК-белкового комплекса с идентификатором 3hdd 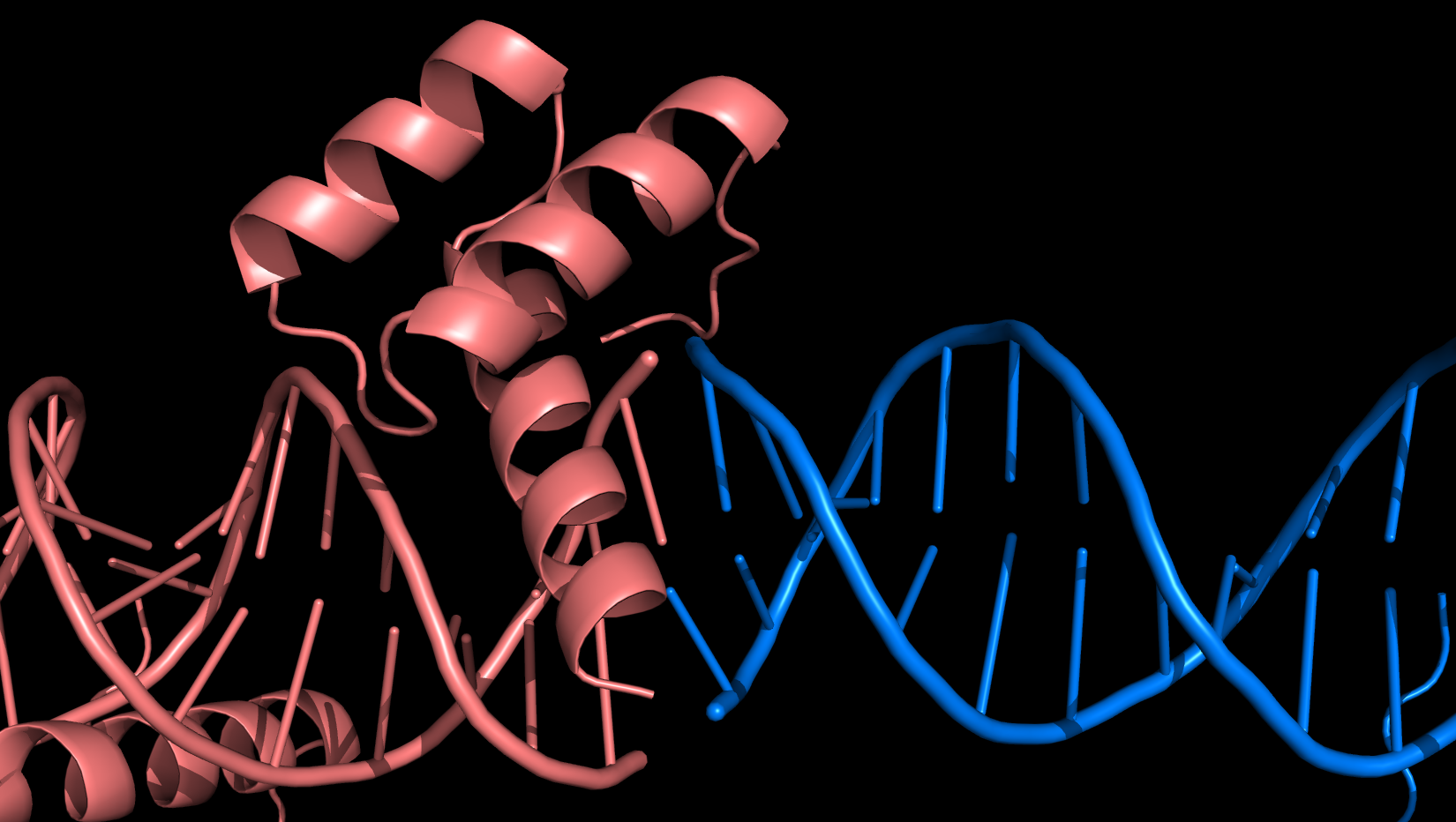 Рисунок 9. Структура ДНК-белкового комплекса с идентификатором 3hdd, 2 соседние кристаллографические ячейки Асимметрическая и биологическая единицыы В рентгеноструктурном эксперименте выделяют асимметрическую единицу - ячейку, движениями которой можно восстановить кристалл. Однако это не всегда совпадает с функциональными особенностями конкретных белков. Часто количество молекул, входящих в асимметрическую единицу не совпадает с теми, которые включены в биологическую. Например, асисмметрической единицей протеазы HhoA бактерии Synechocystis sp (идентификатор PDB - 5B6L) является 1 молекула белка, а функциональный комплекс представляет из себя гомогексамер (рисунки 10 и 11). По одной молекуле можно, и в данном случае удобно, восстановить структуру всего комплекса.
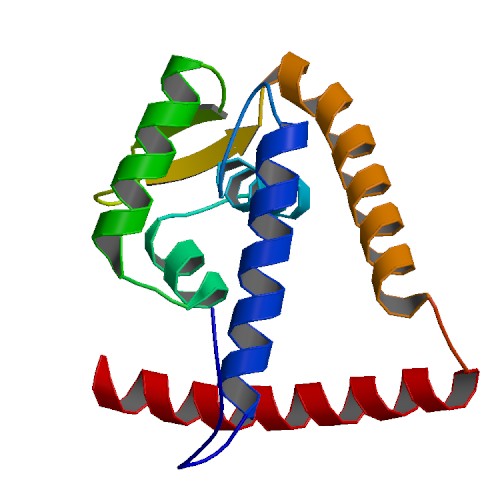
Гексамер - довольно редкий случай, чаще всего белок работает в форме димера. Асимметрической едницей в таком случае может быть, например, димер целиком, как в описанном выше репрессоре транскрипции (идентификатор 3s5r), или нефункциональный мономер, как в случае другого транскрипционного фактора MarR (идентификатор 5eri). Этот белок изображен на рисунках 12-13.
Встречается и обратная ситуация, когда по некоторым причинам исследователям удобнее включить в асимметрическую единицу большее число молекул, чем это есть в настоящем комплексе. |
||||||||||||||||||||||||||||
| © Pogorelskaya Sasha | Last modification date: 29.11.16 | |||||||||||||||||||||||||||||